
高硼土壤增施硫肥对油菜硼吸收与分配的影响
2019-03-29 02:17李鸣凤刘新伟王海彤赵竹青
中国农业科学 2019年5期
李鸣凤,刘新伟,王海彤,赵竹青
高硼土壤增施硫肥对油菜硼吸收与分配的影响
李鸣凤,刘新伟,王海彤,赵竹青
(华中农业大学微量元素研究中心/新型肥料湖北省工程实验中心,武汉 430070)
【目的】探索高硼环境下施用硫肥对油菜硼吸收和分配的影响,为合理施用硫肥缓解油菜硼毒害提供理论基础。【方法】以油菜华油杂9号为试验材料,在高硼灰潮土上(全硼含量13.44 mg·kg-1,有效硼5.03 mg·kg-1)进行盆栽试验。研究了6个硫水平(0、20、50、100、200、500 mg·kg-1)对油菜果荚、叶、茎、根部的硼含量及硼的分配、细胞壁的提取率、细胞壁硼含量和比例的影响。【结果】随着施硫量增加,油菜各部位的生物量呈先增加后降低的趋势,其中施硫50 mg·kg-1时,油菜鲜重生物量最高,可达364.5 g/株。施用不同量硫肥后,油菜硼含量及分配均为果荚>叶>茎>根,表明果荚为硼的主要累积器官。增施硫肥,油菜果荚的硼含量呈下降趋势,施硫量由0增至100 mg·kg-1,油菜果荚硼含量和分配显著降低,降幅分别为14.8%和15.0%。油菜果荚细胞壁提取率增加43.0%,其细胞壁硼含量和比例无显著变化,继续增施硫肥至200 mg·kg-1,油菜果荚硼含量、硼分配比和细胞壁硼均下降至显著水平,且细胞壁硼分配比显著上升。与果荚不同,增施硫肥后油菜叶、茎和根的硼含量则呈缓慢上升的趋势。施硫量由0增至100 mg·kg-1,油菜叶、茎和根硼含量分别增加15.0%、32.9%和34.9%,其分配比例分别增加13.4%、29.6%和18.6%,同时油菜叶片、茎和根部细胞壁硼含量显著增加12.3%、22.9%和14.9%,表明增施硫肥增加了油菜叶、茎和根的硼含量。【结论】硫肥可通过增加油菜生物量,限制硼从根、茎、叶到果荚的转运,以及增加油菜果荚细胞壁硼含量来缓解高硼对油菜果荚的毒害,最佳施硫量为50—100 mg·kg-1。
硼;硫肥;油菜;硼分配;细胞壁
0 引言
【研究意义】硼(B)是植物必需的微量元素之一,适宜植物生长的土壤有效硼浓度范围为0.5—1.0 mg·kg-1[1]。但由于采矿污染、高硼含量灌溉水,以及不平衡肥料施用等原因造成农田土壤中硼积累过高[2]。在高硼土壤上(有效硼>3.0 mg·kg-1)作物会出现不同程度的硼毒害现象[1],例如叶的尖端和边缘表现烧伤和坏死症状[3],根系的生长受到抑制[4],生物量显著下降[5]。目前我国高硼土壤主要集中在硼矿区,如辽宁、吉林、青海和西藏等地区[6]。油菜是十字花科作物,需硼量较大,长江流域属于缺硼地区,油菜种植区域已经推广硼肥施用技术,施肥不当或硼肥逐年累积造成的硼毒害现象时有发生,硼毒害不仅影响油菜的产量和品质,还对动物和人类健康具有潜在的负面影响[7]。且目前部分油菜种植区域忽略了硫肥的施用,出现了高硼缺硫土壤。因此探索油菜种植区域硫缓解硼毒害可行性具有的实际生产意义。【前人研究进展】硫对于油菜是仅次于氮、磷、钾元素之后的第四位元素[8]。适量的硫肥可以促进油菜的生长,提高油菜的产量和品质[9]。研究表明硫元素的吸收和循环显著影响其他元素的吸收和利用,硫与其他元素间的交互作用及其效应要超过它单独的营养作用[10]。硫肥可以增加植物根系中的铜、锌、镉的含量,降低其在植物茎叶中的累积[10]。WANG等研究表明,增施 NaHS,显著降低了硼毒害下的黄瓜幼苗细胞壁果胶甲酯酶(PME)活性,从而减少高硼对根的抑制程度,增强黄瓜幼苗抵抗高硼胁迫的能力[11]。另有研究表明SO42-可能与硼产生互作效益,从而减少植物对硼的吸收利用[12]。目前硫对植物硼吸收利用分配的研究相对较少。【本研究切入点】油菜是世界上第二大食用油来源[13],具有很高的营养价值和良好的脂肪酸组成。作为最大的油菜生产国,中国占全球23.6%的油菜种植面积和占22.2%的全球油菜籽产量[14]。油菜需硫量和需硼量均较大,硫与其他元素的研究多集中在与氮、锌以及重金属等元素的相互关系[10],硫对缓解油菜硼毒害的作用还缺乏系统研究。【拟解决的关键问题】本研究以华油杂9号为试验材料,在高硼缺硫的灰潮土中,通过植物硼吸收分配以及细胞壁硼含量和比例来探索施硫量对油菜硼的吸收利用的影响。
1 材料与方法
1.1 试验土壤和材料
试验土壤为湖北天门油菜田的灰潮土,试验地点为湖北省武汉市华中农业大学。在油菜种植前一周,将过2 mm筛的12 kg土壤与定量基肥拌匀,装于直径为25 cm,高为35 cm的桶中。土样的主要理化性质为pH 7.12、有机质 17.51 g·kg-1、碱解氮 53.12 mg·kg-1、有效磷 5.06 mg·kg-1、速效钾85.03 mg·kg-1、有效硫14.86 mg·kg-1、有效硼5.03 mg·kg-1、全硼13.44 mg·kg-1。
1.2 试验设计
油菜种子在40℃水中浸泡2 h。然后将种子放置在充满超纯水的无菌海绵上催芽,以无菌纱布覆盖,24 h后,选取露白的油菜种子移植在盆栽中,每盆定植1株。试验共6个处理,设6个硫水平:0、20、50、100、200、500 mg·kg-1(表示为S0、S20、S50、S100、S200、S500),4次重复。其他肥料的施肥量为: N 0.3 g·kg-1,K2O 0.32 g·kg-1,P2O50.16 g·kg-1,与土壤混匀。试验于2017年11月种植,2018年3月油菜结荚期(果荚形成期)测定。
1.3 采样与测定
生物量的测定:果荚期油菜分为果荚、叶、茎和根四部分,用去离子洗净,擦干,称量油菜各部位鲜重。
细胞壁的提取及各部位硼含量的测定:提取方法参照 HU等的方法[15],将约3.0000 g的油菜样品用液氮研磨,加入10倍体积的冰水转移至离心管,然后1 000×离心10 min,收集上清液,残渣再用 10 倍体积的冰水冲洗后离心,合并这两次的上清液,测定其中的硼含量作为自由态硼含量(主要指存在于自由空间中的硼);残渣用 10 倍体积80% 乙醇冲洗 3 次,然后用10 倍体积的甲醇-氯仿(体积比1﹕1)混合物冲洗1次,最后用10倍体积丙酮冲洗。上述有机试剂浸提的主要是细胞的原生质体,因此有机试剂提取的这部分硼定义为原生质体硼。最后剩下的不溶性残渣即为粗细胞壁,将其烘干并称重,测定细胞壁硼含量。总硼含量为自由态硼含量,原生质体态硼含量和细胞壁硼含量的总和。姜黄素比色法测定硼含量。
细胞壁提取率(%)=烘干后细胞壁质量/提取细胞壁样品的鲜重×100%;
细胞壁硼含量(mg·kg-1)=粗细胞壁硼含量×细胞壁提取率;
总硼含量(mg·kg-1)=自由态硼含量+原生质体态硼含量+细胞壁硼含量;
细胞壁硼比例(%)=细胞壁硼含量/总硼含量。
叶片的超显微结构测定[16]:取油菜顶4片完全展开叶,自上往下算。用去离子水冲洗干净后,将叶片剪成 1 mm×1 mm左右的小块,快速放入 2.5%的戊二醛固定液中固定 12 h 以上。取出植物材料,用 0.1 mol·L-1的磷酸缓冲液冲洗 4 次,每次 15 min。最后将植物组织依次放入 50%、70%、90%的乙醇中进行脱水 15 min。接着用 90%乙醇和 90%丙酮的等体积混合液脱水15 min,再用 90%的丙酮脱水 15 min,最用在纯丙酮中脱水2次,每次 15 min。整个脱水过程保持组织的温度为 0—4℃。将植物组织用丙酮和包埋液进行包埋和固化处理后,使用超薄切片机将样品组织切成厚度约50—60 nm 的切片,染色后在透射电镜下观察拍照。
1.4 数据分析
本试验采用Excel 2016和SPSS statistics17.0软件进行图表的编辑和试验数据的统计分析处理。
2 结果
2.1 增施硫肥对油菜不同部位生物量的影响
随着施硫量增加,油菜各部位生物量呈现先增加后降低的趋势(表1)。施硫量由0增至50 mg·kg-1,油菜果荚、叶和茎部位生物量显著增加,增幅分别为19.5%、18.0%和17.0%,油菜根部生物量无显著变化,油菜的单株生物量显著增加16.4%。施硫量由50增至500 mg·kg-1,油菜果荚、叶、茎和根的生物量显著降低,降幅分别为15.7%、15.3%、17.6%和13.2%,表明在高硼土壤上施用适量硫肥可促进油菜的生长,但过量的硫肥会抑制油菜的生长。
2.2 增施硫肥对油菜硼含量及分配的影响
不同施硫量下油菜硼含量及其分配均为果荚>叶>茎>根,说明油菜果荚是硼主要累积位点(图1)。随着施硫量增加,油菜果荚硼含量呈显著下降趋势,油菜叶、茎和根硼含量则呈显著上升趋势。施硫量由0增至100 mg·kg-1,油菜果荚硼含量及分配比例显著降低,降幅分别为14.8%和15.0%,然而油菜叶、茎和根硼含量分别增加15.0%、32.9%和34.9%,其分配比例分别增加13.4%、29.6%和18.6%。施硫量由100增至500 mg·kg-1,油菜果荚硼含量和分配比例均显著降低,叶、茎和根硼含量显著增加。表明增施硫肥限制硼从根、茎叶到果荚的移位,增强了油菜对硼的耐受性。

表1 增施硫肥对结荚期油菜不同部位生物量的影响(鲜重FW)
数值为平均值±标准误(=3),同列数值后不同字母代表<0.05 下差异显著。下同
Values are mean ± SE (n = 3), numbers followed by different letters in the same column differ significantly at<0.05. The same as below

图1 增施硫肥对结荚期油菜硼含量(左)及分配(右)的影响
2.3 增施硫肥对油菜细胞壁硼含量和比例的影响
不同施硫量下油菜各部位的细胞壁中硼含量为果荚>叶>茎>根,说明油菜的果荚需硼量较大。随着施硫量的增加,油菜果荚细胞壁硼含量显著下降,相反的是,油菜叶、茎和根细胞壁硼含量显著上升(图2)。施硫量由0增至100 mg·kg-1,油菜果荚胞壁硼含量和比例无显著变化,油菜叶片、茎和根部细胞壁硼含量显著增加,增幅为12.3%、22.9%和14.9%。施硫量由0增至100 mg·kg-1,油菜叶片、茎细胞壁硼比例无显著变化,油菜根部细胞壁硼比例显著下降7.4%。相比不施用硫肥,施硫200 mg·kg-1后,油菜果荚细胞壁硼含量显著下降16.1%,而细胞壁硼分配比例显著上升13.0%。说明增施适量硫肥对油菜果荚细胞壁结合硼含量无显著影响,增施过量硫肥显著降低了油菜细胞壁硼含量。
2.4 增施硫肥对油菜各部位细胞壁提取率的影响
随着施硫量增加,油菜果荚细胞壁提取率呈不断上升趋势。相比不施硫,施硫100 mg·kg-1后,油菜果荚细胞壁提取率增加43.0%(表2)。不同的是随着施硫量的增加,油菜叶片、茎和根部细胞壁提取率呈下降的趋势,施硫量由0增至100 mg·kg-1,油菜叶和茎细胞壁提取率有轻微下降,未达到显著差异,油菜根部细胞壁提取率下降11.0%。相比不施用硫肥,增施硫肥至500 mg·kg-1,油菜叶片、茎和根部细胞壁提取率分别下降17.5%、10.8%和13.7%。表明在高硼土壤上增施硫肥可以提高油菜果荚细胞壁提取率,降低油菜叶、茎和根部细胞壁提取率。

图2 增施硫肥对结荚期油菜细胞壁硼含量(左)和比例(右)的影响
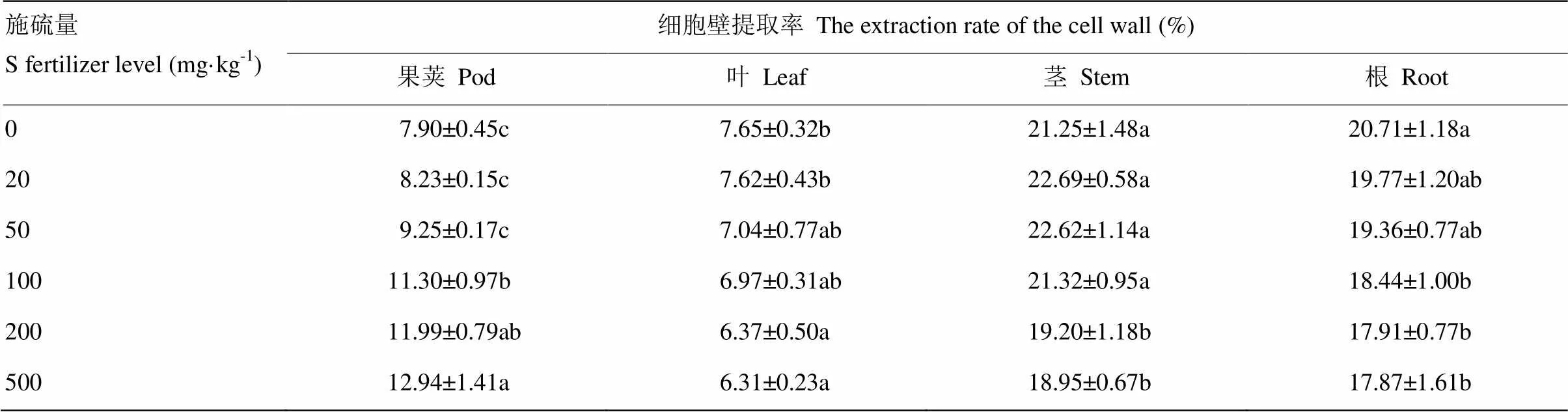
表2 增施硫肥对结荚期油菜各部位细胞壁提取率的影响
2.5 增施硫肥对油菜叶片细胞的超显微结构的影响
在高硼环境下,油菜叶片细胞的叶绿体、线粒体等细胞器等均较为完整(图3)。在缺硫处理下,油菜叶片叶绿体呈细长状,叶绿体和线粒体紧贴细胞壁,细胞壁较厚。施硫100 mg·kg-1后,油菜叶绿体呈饱满梭状,线粒体的脊增加,细胞质较为充盈,细胞壁较薄。施硫500 mg·kg-1后,油菜叶绿体出现变形,轻微解体现象,叶绿体基粒垛叠结构遭到破坏,细胞壁较薄。
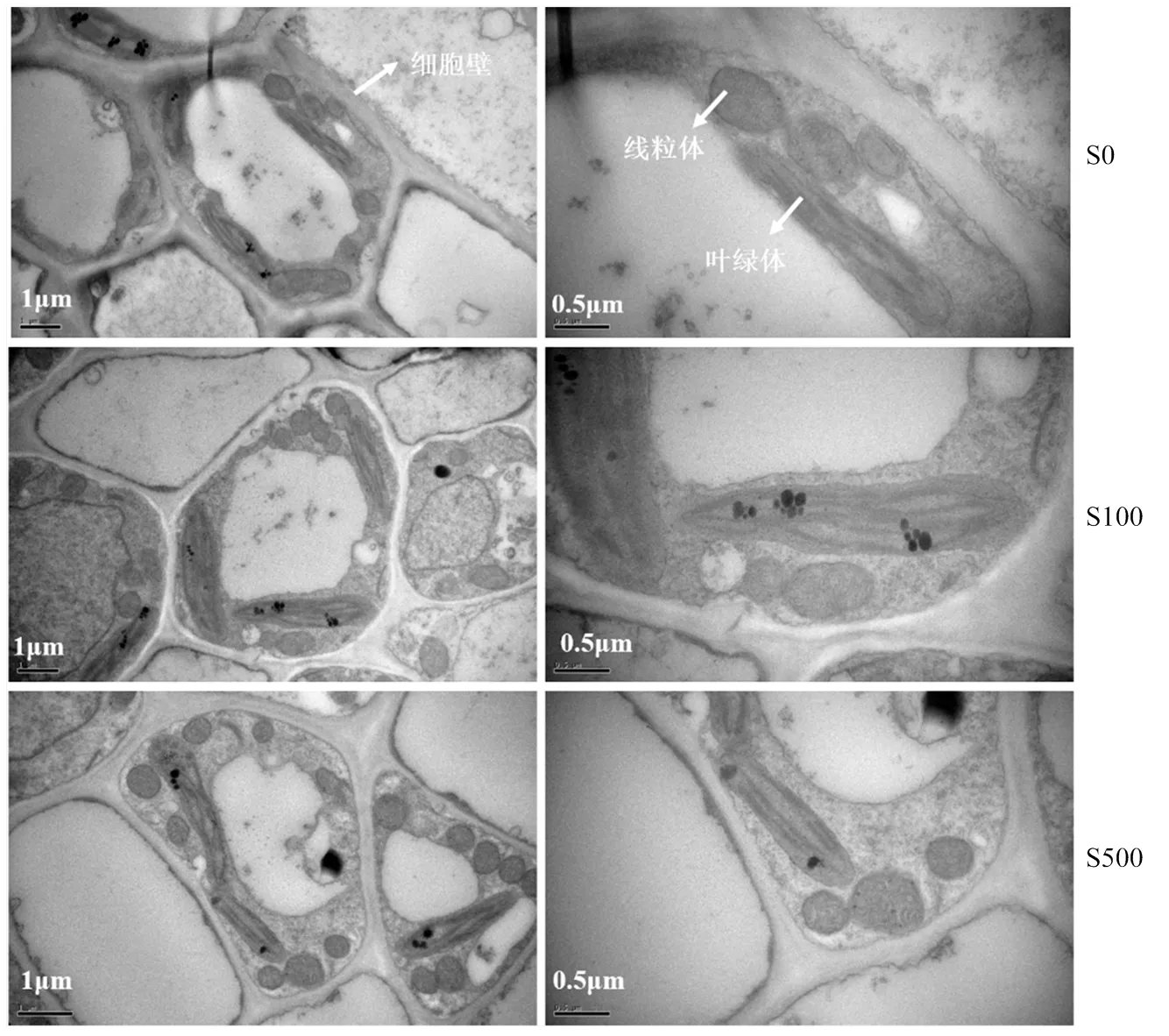
图3 增施硫肥对结荚期油菜叶片透射电镜结构的影响
3 讨论
3.1 高硼土壤上增施适宜硫肥可促进油菜生长发育
油菜对硫的需求量较大,仅次于氮、钾,与磷的需求量较为一致[17]。研究表明适量的增施硫肥,显著促进油菜的生长发育[9]。本试验结果表明随着施硫量的增加,油菜各部位生物量呈现先增加后降低的趋势(图1)。施硫量小于50 mg·kg-1,显著提高了油菜生物量。继续增施硫肥100 mg·kg-1时油菜生物量无显著差异,施硫量高于100 mg·kg-1时,油菜各部位生物量显著下降,原因在于试验土壤为低硫土壤(有效硫为14.86 mg·kg-1),且油菜需硫量较大,因而适宜的增施硫肥促进了油菜的生长,继续增施硫肥高于100 mg·kg-1,大量的硫肥可能影响土壤的pH[18-19]、微生物[20]或直接导致植物硫中毒[21]等,从而抑制了植物的生长。油菜叶片的超显微结构表明在施硫量为100 mg·kg-1油菜叶片的叶绿体、线粒体等器官结构最为完整(图3),有利于油菜光合作用,从而促进生物量的累积。总之,在高硼灰潮土上,硫对于油菜生物量表现为低浓度促进,高浓度抑制,适宜的施硫量为50—100 mg·kg-1。
3.2 高硼土壤上增施硫肥可缓解油菜果荚硼毒害
研究表明当环境中硼浓度较低时,植物不同部位的硼重新活化,优先供应植物的生殖结构,但是,当环境中硼浓度较高时硼主要累积在具有高蒸腾速率的器官叶片中[22]。油菜不同于其他作物,果荚期油菜叶片慢慢凋零,角果既是油菜重要的光合器官又是经济器官,油菜籽粒近2/3的产量来自角果皮的光合作用[23]。果荚期油菜各部位硼含量及其分配均为果荚>叶>茎>根,说明果荚期油菜的果荚是硼主要累积位点,更容易受到过量硼的伤害(图1)。SAVIĆ等[24]研究表明油菜地上部组织浓度约高于10 mg·kg-1FW时,说明油菜处于硼毒害范围。而本试验中不施硫处理,油菜果荚、叶片、茎和根硼浓度分别为33.64、10.02、3.89和3.11 mg·kg-1(图1),表明油菜遭受了硼毒害的部位主要为果荚。施硫量由0增至100 mg·kg-1油菜果荚硼含量显著下降14.8%,然而其他部位硼含量显著上升(图1),适量的增施硫肥主要是缓解高硼对油菜果荚伤害。REID[25]研究表明对作物对硼的耐受性与减少硼向叶中运输和积累的能力有关。本试验结果表明在高硼土壤上增施硫肥,油菜果荚硼的分配显著下降,而叶、茎和根的硼分配比例显著上升,表明增施硫肥不仅通过生物量的稀释效应来缓解油菜硼毒害还通过限制硼从根茎叶向果荚的转移,从而增强了油菜对硼的耐受性。
3.3 高硼土壤上增施硫对油菜细胞壁的影响
硼主要作用于细胞壁,有研究表明细胞中的硼 80%以上存在于细胞壁中,硼对细胞壁的结构和功能有重要影响[26]。硼在植物细胞壁的含量和相对分布取决于硼的供应,在硼供应相对多的环境,植物自由态硼、原生质体态硼以及细胞壁硼含量均高,细胞壁硼占总硼的比例相对较低[27]。本试验中在高硼土壤上,油菜各部位细胞壁硼比例为16.3%—37.0%(图2)。相比不施用硫肥,施硫100 mg·kg-1,油菜果荚总硼含量显著降低14.8%,而果荚细胞壁硼含量和比例无显著影响,主要原因在于细胞壁具有有限的硼结合位点[28],且增施硫肥主要降低了油菜自由态硼或原生质体态硼的含量。施硫量由0增至100 mg·kg-1,油菜叶片、茎和根部细胞壁硼含量显著增加12.3%、22.9%和14.9%,由此表明油菜叶片、茎和根部细胞壁硼的结合位点并没有与硼完全结合,表明增施硫肥至100 mg·kg-1,油菜叶片、茎和根部硼含量增加并没有使其处于硼毒害水平。相比不施用硫肥,施硫200 mg·kg-1,油菜果荚硼含量和细胞壁硼含量均显著下降,细胞壁硼比例显著上升,表明施硫量高于200 mg·kg-1后,油菜果荚细胞壁硼含量已经下降至正常水平。
研究表明硼与RG-II的交联增加了细胞壁的机械强度并保证了膨压驱动的细胞的正常生长[29]。硼在细胞壁上的结合与细胞壁孔隙大小密切相关,细胞壁孔隙大小直接控制着离子和小分子有机物在细胞中的自由渗透扩散[30]。本试验的结果表明,在高硼土壤上增施硫肥,油菜果荚的硼含量下降,细胞壁提取率增加,表明油菜可能是通过增加细胞壁厚度,降低硼等小分子进入油菜果荚细胞,从而缓解硼毒害对油菜果荚的伤害。
4 结论
高硼环境下,果荚期的油菜各部位中果荚为硼的主要累积位点,更容易受到过量硼伤害。适量硫肥通过增加油菜生物量,限制硼从根、茎、叶转运到果荚,降低了油菜果荚硼含量和分配比例,从而提高油菜硼的耐受能力。同时,施硫可通过增加细胞壁提取率、降低果荚细胞壁硼含量,缓解过量硼对油菜果荚的伤害。在高硼环境下,油菜适宜的施硫量为50—100 mg·kg-1。
[1] CARTWRIGHT B, ZARCINAS B A, MAYFIELD A H. Toxic concentration of boron in a red-brown earth at Gladstone South Australia., 1984, 22(3): 261-272.
[2] PARKS J L, EDWARDS M. Boron in the Environment., 2005, 35(2): 81-114.
[3] GÜR N, TÜRKER O C, BÖCÜK H. Toxicity assessment of boron (B) byL. andL. and their possible use as model plants for ecological risk assessment of aquatic ecosystems with boron pollution., 2016, 157: 1-9.
[4] SHAH A, WU X W, ULLAH A, FAHAD S, MUHAMMAD R, YAN L, JIANG C C. Deficiency and toxicity of boron: alterations in growth, oxidative damage and uptake by citrange orange plants., 2017, 145(6): 575-582.
[5] 段碧辉, 刘新伟, 矫威, 赵竹青, 胡承孝. 硒减轻油菜幼苗硼毒害机理的研究. 中国农业科学, 2014, 47(11): 2126-2134.
DUAN B H, LIU X W, JIAO W, ZHAO Z Q, HU C X. Alleviation of boron toxicity on rape seedlings by selenium., 2014, 47(11): 2126-2134. (in Chinese)
[6] 王春利, 邢小茹, 吴国平, 司杨, 魏复盛. 宽甸土壤及部分农作物中硼的分布及污染分析. 中国环境监测, 2003, 19(5): 4-7.
WANG C L, XING X R, WU G P, SI Y, WEIF S. Analyses of boron distribution and pollution in soil and some crops in Kuandian., 2003, 19(5): 4-7. (in Chinese)
[7] SCIALLI A R, BONDE J P, BRÜSKE-HOHLFELD I, CULVER B D, Li Y, SULLIVAN F M. An overview of male reproductive studies of boron with an emphasis on studies of highly exposed Chinese workers., 2010, 29(1): 10-24.
[8] 黄启为, 杨志辉, 胡树林,刘鹏, 王翠红, 彭建伟.硫肥对油菜产量及品质的影响. 湖南农业大学学报(自然科学版), 2001, 27(4): 276-279.
HUANG Q W, YANG Z H, HU S L, LIU P, WANG C H, PENG J W. Effects of sulfur fertilizer on yield and quality of rapeseed., 2001, 27(4): 276-279. (in Chinese)
[9] 吴曦, 陈明昌, 杨治平. 碱性土壤施硫磺对油菜生长、土壤pH和有效磷含量的影响. 植物营养与肥料学报, 2007, 13(4): 671-677.
WU X, CHEN M C, YANG Z P. Effects of sulfur application on the growth of cole, soil pH and available P in alkaline soil., 2007, 13(4): 671-677. (in Chinese)
[10] 孟赐福, 姜培, 曹志洪, 徐秋芳, 周国模. 硫素与其他营养元素的交互作用对作物养分吸收、产量和质量的影响. 土壤, 2009, 41(3): 329-334.
MENF C F, JIANG P K, CAO Z H, XU Q F, ZHOU G M. Interaction effects of sulfur and other nutrients on nutrient uptakes, yields, and qualities of crops., 2009, 41(3): 329-334. (in Chinese)
[11] WANG B L, SHI L, LI Y X, ZHANG W H. Boron toxicity is alleviated by hydrogen sulfide in cucumber (L.) seedlings., 2010, 231(6): 1301-1309.
[12] SMITH T E, GRATTAN S R, GRIEVE C M, POSS J A, SUAREZ D L. Salinity's influence on boron toxicity in broccoli: II. Impacts on boron uptake, uptake mechanisms and tissue ion relations., 2010, 97(6): 783-791.
[13] FOLEY J A, RAMANKUTTY N, BRAUMAN K A, CASSIDY E S, GERBER J S, JOHNSTON M, MUELLER N D, O'CONNELL C, RAY D K, WEST P C, BALZER C, BENNETT E M, CARPENTER S R, HILL J, MONFREDA C, POLASKY S, ROCKSTRÖM J, SHEEHAN J, SIEBERT S, TILMAN D, ZAKS D P. Solutions for a cultivated planet., 2011, 478(7369): 337-342.
[14] FAO. Food and agriculture organization of the united nations, 2016. http://www.fao.org/faostat/en/#data/QC.
[15] HU H, BROWN P H. Localization of boron in cell walls of squash and tobacco and its association with pectin evidence for a structural role of boron in the cell wall., 1994, 105(2): 681-689.
[16] 刘磊超, 姜存仓, 董肖, 吴秀文, 刘桂东, 卢晓佩. 硼胁迫对枳橙砧木细根根尖成熟区和幼嫩叶片细胞结构的影响. 中国农业科学, 2015, 48(24): 4957-4964.
LIU L C, JIANG C C, DONG X, WU X W, LIU G D, LU X P. Effects of boron deficiency on cellular structures of maturation zone from root tips and functional leaves from middle and upper plant in trifoliate orange rootstock., 2015, 48(24): 4957-4964. (in Chinese)
[17] WEIL R R, MUGHOGHO S K. Sulfur nutrition of maize in four regions of Malawi., 2000, 92(4): 649.
[18] 刘新伟, 段碧辉, 夏全杰, 矫威, 郭再华, 胡承孝, 赵竹青. 硫对土壤中硒形态变化及油菜硒吸收的影响. 环境科学, 2014, 35(9): 3564-3571.
LIU X W, DUAN B H, XIA Q J, JIAO W, GUO Z H, HU C X, ZHAO Z Q. Effects of sulfur on transformation of selenium in soil and uptake of selenium in rape., 2014, 35(9): 3564-3571. (in Chinese)
[19] 刘新伟, 段碧辉, 赵小虎, 郭再华, 胡承孝, 赵竹青.外源四价硒条件下硫对小麦硒吸收的影响机制. 中国农业科学, 2015, 48(2): 241-250.
LIU X W, DUAN B H, ZHAO X H, GUO Z H, HU C X, ZHAO Z Q. Effects of sulfur on selenium uptake in wheat and its mechanism when amended with selenite., 2015, 48(2): 241-250. (in Chinese)
[20] 谢晓梅, 廖敏, 张楠, 徐培智, 徐昌旭, 刘光荣. 外源S2-抑制水稻生长及土壤微生物活性的半效应浓度研究. 植物营养与肥料学报, 2015, 21(5): 1286-1293.
XIE X M, LIAO M, ZHANG N, XU P, XU CX, LIU G R. EC50of soil S2-inhibiting rice growth and soil microbial activities.2015, 21(5): 1286-1293. (in Chinese)
[21] 董丽欣, 李保国, 齐国辉, 郭素萍, 张雪梅,于祎飞, 胡志伟. 土壤铜、硫污染对苹果幼树生长发育的影响. 水土保持学报, 2011, 25(6): 198-201.
DONG L X, LI B G, QI G H, GUO S P, ZHANG X M, YU W F, HU Z W. Effects of soil copper and sulfur pollution on growth of young apple trees., 2011, 25(6): 198-201. (in Chinese)
[22] BOGIANI J C, SAMPAIO T F, ABREU-JUNIOR C H, ROSOLEM C A. Boron uptake and translocation in cotton cultivars., 2013, 375(1/2), 241-253.
[23] 冷锁虎, 唐瑶, 李秋兰, 左青松, 杨萍. 油菜的源库关系研究Ⅰ.角果大小对油菜后期源库的调节. 中国油料作物学报, 2005, 27(3): 37-40.
LENG S H, TANG Y, LI Q L, ZUO Q S, YANG P. Studies on source and sink of rapeseed.Ⅰ. Regulation of pod size on source and s ink in rapeseed after flowering., 2005, 27(3): 37-40. (in Chinese)
[24] SAVIĆ J, MARJANOVIĆ-JEROMELA A, GLAMOČLIJA D, PRODANOVIĆ S. Oilseed rape genotypes response to boron toxicity., 2013, 45(2): 565-574.
[25] REID R. Update on boron toxicity and tolerance in plants. Advances in plant and animal boron nutrition., Dordrecht, The Netherlands, 2007: 83-90.
[26] MATOH T, ISHIGAKI K, OHNO K, AZUMA J. Isolation and characterization of a boron-polysaccharide complex from radish roots., 1993, 34(4): 639-642.
[27] 刘桂东, 胡萍, 张婧卉, 周高峰, 曾钰, 钟八莲, 姜存仓. 缺硼对脐橙幼苗硼分配及叶片细胞壁组分硼含量的影响. 植物营养与肥料学报, 2018, 24(1): 179-186.
LIU G D, HU P, ZHANG J H, ZHOU G F, ZENG Y, ZHONG B L, JIANG C C. Effect of boron deficiency on boron distribution in different plant parts and boron concentration in leaf cell wall components in navel orange plants., 2018, 24(1): 179-186. (in Chinese)
[28] DANNEL F, PFEFFER H, RÖMHELD V. Compartmentation of boron in roots and leaves of sunflower as affected by boron supply., 1998, 153(5/6): 615-622.
[29] O’NEILL M A, EBERHARD S, ALBERSHEIM P, DARVILL A G. Requirement of borate cross-linking of cell wall rhamnogalacturonan II for Arabidopsis growth., 2001, 294(5543): 846-849.
[30] 李金柱, 吴礼树, 杨玉华. 硼在植物细胞壁上营养机理的研究进展. 中国油料作物学报, 2004, 26(4): 96-100.
LI J Z, WU L S, YANG Y H. Advances on the study of B nutritional mechanisms in plant cell wall., 2004, 26(4): 96-100. (in Chinese)
Effects of Sulfur Fertilizer on Boron Uptake and Distribution of Rape in B-contaminated Soil
LI MingFeng, LIU XinWei, WANG HaiTong, ZHAO ZhuQing
(Microelement Research Center, Huazhong Agricultural University/Hubei Provincial Engineering Laboratory for New-Type Fertilizer, Wuhan 430070)
【Objective】This study explored the effects of sulfur (S) fertilizer application on B uptake and distribution in rapeseed under high boron (B) environment, and provided a theoretical basis for rational application of S fertilizer to alleviate B toxicity in rapeseed (L.). 【Method】Rape plants (cv. Huayouza 9) were grown in pots filled with calcareous alluvial soil contaminated with B (total B content: 13.44 mg·kg-1; available B: 5.07 mg·kg-1) and the effects of S fertilizer level (0, 20, 50, 100, 200, and 500 mg·kg-1) on B concentration, distribution, cell wall extraction rate, cell wall B concentration and its proportion of total B in different parts of the rape were determined at the podding stage. 【Result】 With the increase of S application, the biomass increased at first and then decreased in different parts of rape. When the S was applied at 50 mg·kg-1, the fresh biomass of rape was the largest, reached 364.5 g/plant under different S application, and the B concentration and distribution in different parts of rape followed the order of pods>leaves>stems>roots, which indicated that the pods were the main accumulation site of B. As the S fertilizer level increased from 0 to 100 mg·kg-1, the pod B concentration and distribution ratios decreased by 14.8% and 15.0%, respectively. The extraction rate of cell wall of rapeseed pods increased by 43.0%, and there was no significant change in the B concentration and proportion in the cell wall. When S fertilizer was continuously applied to 200 mg·kg-1, the Bconcentration, distribution ratios and cell wall B concentration of rapeseed pods decreased significantly, and cell wall B proportion increased significantly compared with that of non-sulfur fertilizer. The opposite trends were found in the B concentrations and cumulative distribution ratios in the leaves, stems and roots. As the S fertilizer level increased from 0 to 100 mg·kg-1, the B concentration significantly increased by 15.0%, 32.9% and 34.9% in rape leaves, stems, and roots, respectively. The B distribution ratios significantly increased by 13.4%, 29.6% and 18.6%, and B concentration in the cell wall significantly increased by 12.3%, 22.9% and 14.9% in rape leaves, stems, and roots, respectively. It showed that the addition of S fertilizer increased the B concentration of the leaves, stems and roots of rapeseed. 【Conclusion】S fertilizer application effectively alleviated B toxicity by increasing biomass, limiting B translocation from roots, stems and leaves to pods, and by increasing the extraction rate of cell wall in rape plants grown on B-contaminated soil. The optimum amount of S fertilizer application was 50-100 mg·kg-1.
boron contamination; sulfur fertilizer; rape; boron distribution; cell wall
10.3864/j.issn.0578-1752.2019.05.009
2018-09-02;
2018-12-17
中央高校基本科研业务费专项基金(2662018PY002)
李鸣凤,Tel:13554313504;E-mail:mingfengL0124@163.com。通信作者赵竹青,E-mail:zzq@mail.hzau.edu.cn
(责任编辑 李云霞)
猜你喜欢
中国真菌学杂志(2022年1期)2023-01-06
中国交通信息化(2022年7期)2022-10-27
福建林业科技(2020年4期)2020-12-31
今日农业(2020年19期)2020-12-14
三农资讯半月报(2020年18期)2020-10-14
科学导报·学术(2020年19期)2020-07-09
安徽农学通报(2020年7期)2020-05-26
农民致富之友(2019年14期)2019-05-24
数码世界(2018年1期)2018-12-23
江苏农业科学(2016年11期)2017-03-21
