
非标记定量蛋白质组方法分析鲟鱼肽抗D-半乳糖导致的小鼠衰老作用的研究
2019-03-28 07:23:58,,,,*,,*
食品工业科技 2019年3期
,,, ,*,,*
(1.大连医科大学附属第二医院临床营养科,辽宁大连 116023; 2.大连大学附属中山医院 麻醉二科,辽宁大连 116001; 3.大连海洋大学食品科学与工程学院,辽宁大连 116023)
鲟鱼类为一类软骨硬鳞鱼,具有极高的经济价值。中国现有鲟鱼类为2科3属8种,分布于长江水系、黑龙江水系以及新疆[1]。鲟鱼不仅有丰富的营养,还拥有重要的药用价值。据《本草拾遗》记载[2]:“鲟鱼,肉味甘,性平,无毒。补虚益气,令人肥健。”。研究表明,鲟鱼含有丰富的不饱和脂肪酸、硫酸软骨素、蛋白质、矿物质元素等[3]。这些物质往往具有多种多样的生理活性。因此,鲟鱼具有潜在的抗氧化、抗衰老、抗疲劳作用。寻找鲟鱼的潜在药用活性部位,是利用好鲟鱼资源的关键之一。研究发现,鲟鱼头的软骨中含有大量的硫酸软骨素,因此鲟鱼头也成为硫酸软骨素新的来源。鲟鱼头经过酶解等加工过程,可以获得肽类、多糖类等具有利用价值的化合物,不但可以获得鲟鱼药用部位的研究结果,而且可以极大地提高鲟鱼产业的经济效益。
衰老是机体各组织器官在多种原因、多种生理反应作用下发生的结果。在提出的许多衰老原因中,免疫功能下降学说和自由基学说具有一定的地位[4]。近年来,开发具有抗衰老作用的多肽类物质成为热点。樊金娟等[5]研究了经中性蛋白酶酶解米糠所获得的米糠抗氧化肽的抗衰老作用,先利用D-半乳糖致衰小鼠构建模型,再灌胃米糠抗氧化肽,结果表明,米糠抗氧化肽组与衰老组相比,高剂量(500 mg/kg bw)米糠抗氧化肽可显著提高致衰老小鼠心脑线粒体过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-Px)、琥珀酸脱氢酶(SDH)和总三磷酸腺苷酶(T-ATPase)活性(p<0.05),明显降低小鼠脑线粒体DNA(mtDNA)缺失突变水平(p<0.01),效果优于中、低剂量米糠抗氧化肽组,米糠抗氧化肽具有较好的抗衰老作用。吉雅等[6]研究了大豆短肽颗粒剂对衰老小鼠模型的作用,发现大豆短肽可以显著降低衰老小鼠血清和肝组织中的丙二醛(MDA)含量,并显著升高其超氧化物歧化酶(SOD)活力,而且大豆短肽还可以显著提高小鼠的免疫功能,促进其脾细胞增殖,因此大豆短肽颗粒剂有显著的抗衰老和增强免疫的作用。曹新志[7]研究了富含谷胱甘肽制品对小鼠抗衰老能力的影响,发现饲喂富含谷胱甘肽制品的小白鼠显著提高了其抗衰老能力。李琳等[8]利用D-乳糖致衰小鼠模型灌胃鳙鱼肽,发现其可使致衰小鼠肝脑组织中MDA的含量明显下降,脑组织中单胺氧化酶(MAO-B)的活性显著降低(p<0.01),抗氧化酶SOD和谷胱甘肽过氧化物酶的活性有所提高,并能使致衰小鼠吞噬细胞的吞噬能力显著提高(p<0.01)。
鲟鱼养殖目前成为水产养殖的一个热点。养殖鲟鱼的主要目的是为了获得鲟鱼卵,它是高级鱼子酱的重要原料。鲟鱼头是鲟鱼加工的主要副产物,目前鲟鱼头并没有得到很好的利用,主要是因为鲟鱼肽(Acipenser schrenki Brandt peptides,ASP)的生物活性没有很好的研究。本论文对从鲟鱼副产物制备的ASP的性质进行研究,进而利用D-半乳糖致衰老小鼠模型以及非标记定量蛋白质组学技术研究鲟鱼肽的抗衰老作用,以期从分子水平上阐述鲟鱼肽抗衰老机理。
1 材料与方法
1.1 材料与仪器
鲟鱼肽(Acipenser schrenki Brandt peptides,ASP) 分子量介于805~3300 Da,长阳清江鹏博开发有限公司(湖北武汉);SPF级雄性昆明小鼠(体重18~22 g) 大连医科大学实验动物中心[SCXK(辽)2008-0002];总抗氧化能力(T-AOC)检测试剂盒、SOD测定试剂盒、MDA测定试剂盒、考马斯亮蓝蛋白质测定试剂盒 南京建成生物工程公司;D-半乳糖、十二烷基硫酸钠(SDS)、尿素、二硫苏糖醇(DTT)、吲哚乙酸(IAA) Bio-Rad公司;三羟甲基氨基甲烷(Tris)、NH4HCO3、三氟乙酸(TFA)、C18Cartridge Sigma公司;BCA定量试剂盒 上海碧云天生物技术有限公司;胰蛋白酶(Trypsin) Promega公司;10 kDa超滤离心管 Sartorius公司;小鼠-3多重亲和去除液相色谱柱(Multiple Affinity Removal LC Column-Mouse 3)、上样柱(Thermo Scientific Acclaim PepMap100,100 μm×2 cm,nanoViper C18)、分析柱(Thermo scientific EASY column,10 cm,ID75 μm,3 μm,C18-A2) Agilent公司;其它试剂 为国产分析纯。
5430R低温高速离心机、真空离心浓缩仪 Eppendordf公司;Triple TOF 6600+质谱仪 AB SCIEX公司;Agilent 1290 Infinity LC超高压液相色谱仪 Agilent公司。
1.2 D-半乳糖致小鼠衰老模型的建立、给药和处理
将60只SPF级昆明小鼠(雄性)称重后随机分为5组,分别为正常组、衰老模型组、鲟鱼肽低、中、高浓度剂量组,其中除正常组每天注射等体积生理盐水外,其他各组每天都皮下组织注射0.5 mL 5% D-半乳糖(生理盐水配制)。低、中、高浓度剂量组每日分别灌胃一次低浓度100 mg/(kg·d)、中浓度200 mg/(kg·d)、高浓度400 mg/(kg·d)鲟鱼肽水溶液,正常组和衰老模型组给予生理盐水(0.1 mL/20 g),连续灌胃6周后,称重,摘眼球取血,离心(3000 r/min 10 min),取上层血清,-20 ℃冰箱冻存备用。处死小鼠后迅速取其肝脏和大脑,生理盐水洗净,用滤纸吸干血水,各组织称取0.3 g,制成10%匀浆液,离心(4500 r/min 10 min),取上清液,4 ℃备用。
1.3 小鼠体质量及组织脏器指数测定
小鼠在电子天平上称重后处死,快速取出肝脏、脑、胸腺和脾,生理盐水洗净,用滤纸吸干血水,直接在电子天平上称重,脏器指数=脏器质量(mg)/小鼠体质量(g)。
1.4 生化指标测定
按试剂盒说明测定小鼠血清中T-AOC能力、SOD活性和MDA含量。
1.5 蛋白质组样品制备
取D-半乳糖致小鼠衰老模型组小鼠血清和鲟鱼肽灌胃中、高剂量组等比例混合小鼠血清各50 μL,均采用去血清高丰度亲和色谱柱,按照Agilent对应方案[9-11]中的操作方法获得小鼠血清中低丰度蛋白组分。经超滤浓缩后加入等体积SDT裂解液,沸水浴裂解小鼠血清中低丰度蛋白15 min,再于14000×g条件下离心20 min。上清采用BCA法进行定量。鲟鱼肽灌胃小鼠血清低丰度蛋白质样品标记为Z,D-半乳糖致小鼠衰老模型组小鼠血清低丰度蛋白质样品标记为M。
1.6 SDS-PAGE电泳
取未经去血清高丰度亲和色谱柱处理的模型组小鼠血清蛋白(Ori)、D-半乳糖致小鼠衰老模型组小鼠血清低丰度蛋白质的3个平行样本(分别标记为M1、M2、M3)和鲟鱼肽灌胃小鼠血清低丰度蛋白质的3个平行给药综合组样品(分别标记为Z1、Z2、Z3)各20 μg,按5∶1(体积比)加入5倍上样缓冲液,沸水浴5 min,进行12.5% SDS-PAGE电泳。条件:恒流15 mA,电泳时间60 min,考马斯亮蓝染色。
1.7 样品酶解
分别取各组蛋白质样品30 μL,加入0.003 mmol DTT,沸水浴5 min后冷却至室温。加入200 μL尿素缓冲液(8 mol/L尿素,150 mmol/L Tris-HCl,pH8.0)混匀后,在10 kDa超滤离心管中于14000×g离心15 min,弃滤液(重复该步骤一次:加尿素缓冲液,混匀,超滤),加入100 μL 100 mmol/L IAA缓冲液(以尿素缓冲液溶解),在600 r/min条件下振荡1 min,室温避光30 min后于14000×g离心15 min,加入100 μL尿素缓冲液,于14000×g离心15 min(该步骤重复两次:加IAA缓冲液、振荡、避光、离心、加尿素缓冲液、离心),加入100 μL 25 mmol/L NH4HCO3溶液,于14000×g离心15 min两次,加入40 μL 0.1 g/L胰蛋白酶缓冲液(以100 mmol/L NH4HCO3溶液配制),600 r/min振荡反应1 min,37 ℃静置16 h。换新收集管,于14000×g离心15 min,再加入40 μL 25 mmol/L NH4HCO3,于14000×g离心15 min并收集滤液。通过C18Cartridge脱盐冻干后,以40 μL 0.1%甲酸溶液复溶,测定280 nm吸光值。
1.8 酶解产物的LC-MS/MS鉴定
酶解后的样品,采用纳升流速的HPLC液相系统Easy nLC进行分离。A相:0.1%甲酸水溶液,B相:0.1%甲酸乙腈水溶液(乙腈为84%)。色谱柱以300 nL/min 95%的A相平衡,样品依次通过上样柱和分析柱分离。0~50 min,B相线性梯度0%~35%;50~55 min,B相线性梯度35%~100%;55~60 min,B相维持100%。经色谱分离后的样品用Q-Exactive质谱仪进行质谱分析。
1.9 MaxQuant的数据分析
质谱分析原始数据为RAW文件,采用MaxQuant软件(版本号1.3.0.5)[12-13]进行查库鉴定,并根据Label free算法进行定量分析[14-15]。
1.10 基因本体(Gene Ontology,GO)功能注释
利用Blast2GO[16]对目标蛋白质集合进行GO注释。利用NCBI BLAST+(ncbi-blast-2.2.28+-win32.exe)比对小鼠血清数据库,保留E值≤0.001的前10条,再依次通过提取、注释。
1.11 KEGG通路注释及其注释的富集分析
利用KEGG Automatic Annotation Server软件[17]获取目标蛋白质序列参与的通路信息。通过Fisher精确检验评价GO term或KEGG通路蛋白质富集度的显著性水平。
1.12 数据处理
2 结果与分析
2.1 衰老模型组动物一般情况
衰老模型组的小鼠连续6周皮下组织注射5% D-半乳糖0.5 mL/d(生理盐水配制)后,小鼠毛色变灰暗,失去光泽,形体瘦弱,行动缓慢以及精神萎靡不振,体质量仅增加18.02 g,增加量明显低于正常级(表1),呈现出明显衰老体征,表明建模成功。

表1 对小鼠体质情况及脏器指数的影响Table 1 Effects of ASP on body weight and its organs index of mice
2.2 治疗组小鼠体质变化情况及脏器指数
各组SPF级雄性昆明小鼠灌胃对应溶液6周后,进行体重变化及脏器指数统计(表1),结果表明,各组小鼠体重均有所增加,但增加量各不相同。正常组小鼠体质量增加21.83 g,鲟鱼肽低、高剂量组体重增加量分别为17.22、18.07 g,与正常组相比具有显著差异(p<0.05),中剂量组增加量为20.04 g,与正常组相比无显著性差异(p>0.05)。D-半乳糖致小鼠衰老时,致使小鼠代谢紊乱,进而导致体重增加缓慢[18]。
在脏器指数中,各给药组的肝指数和胸腺指数与正常组相比不显著(p>0.05),但中、高剂量组的脾指数与正常组相比,增加显著(p<0.05),低剂量组的脾指数增加不显著(p>0.05);中剂量组的脑指数低于正常组,变化显著(p<0.05),且与模型组相比,差异极显著(p<0.01),而低、高剂量组的脑指数与正常组相比,差异不显著(p>0.05)。脾脏是重要的免疫器官,灌胃鲟鱼肽后,可以增加脾脏指数,可在一定程度上增强小鼠机体免疫功能。
2.3 生化指标检测
对小鼠血清中T-AOC、SOD活性和MDA含量进行测定,结果见表2。由表2可见,与模型组相比,灌胃鲟鱼肽中、高剂量组极显著升高了小鼠血清中T-AOC和SOD活性(p<0.01)。灌胃鲟鱼肽低、中、高剂量组均能极显著降低MDA含量(p<0.01)。注射D-半乳糖后,小鼠体内T-AOC下降,SOD活性降低,MDA含量升高,从而引起小鼠衰老现象发生,灌胃鲟鱼肽后,小鼠体内T-AOC增加,因此,鲟鱼肽具有抗D-半乳糖致小鼠衰老作用。
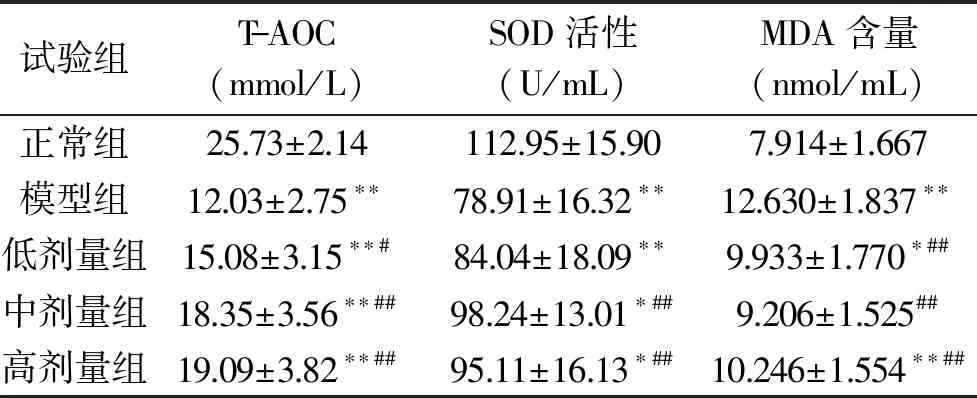
表2 小鼠血清中T-AOC、SOD活性和MDA含量Table 2 T-AOC,SOD activity and MDA content in serum in mice
2.4 血清样品SDS-PAGE和质谱测定
SDS-PAGE(见图1A)结果表明,未经去血清高丰度亲和色谱柱处理的模型组小鼠血清蛋白(Ori)中66 kDa附近蛋白含量过高,影响低丰度蛋白分析,而D-半乳糖致小鼠衰老模型组小鼠血清低丰度蛋白质的3个平行样本M1、M2、M3和鲟鱼肽灌胃小鼠血清低丰度蛋白质的3个平行给药综合组样品Z1、Z2、Z3电泳条带清晰,且各平行组内样品平行性良好,可以进行质谱验证样品平行性。质谱分析Basepeak结果见图1B和图1C,从Basepeak图谱可以看出,组间样本平行性好,表明酶解效果良好,适合进行非标记定量质谱测定。
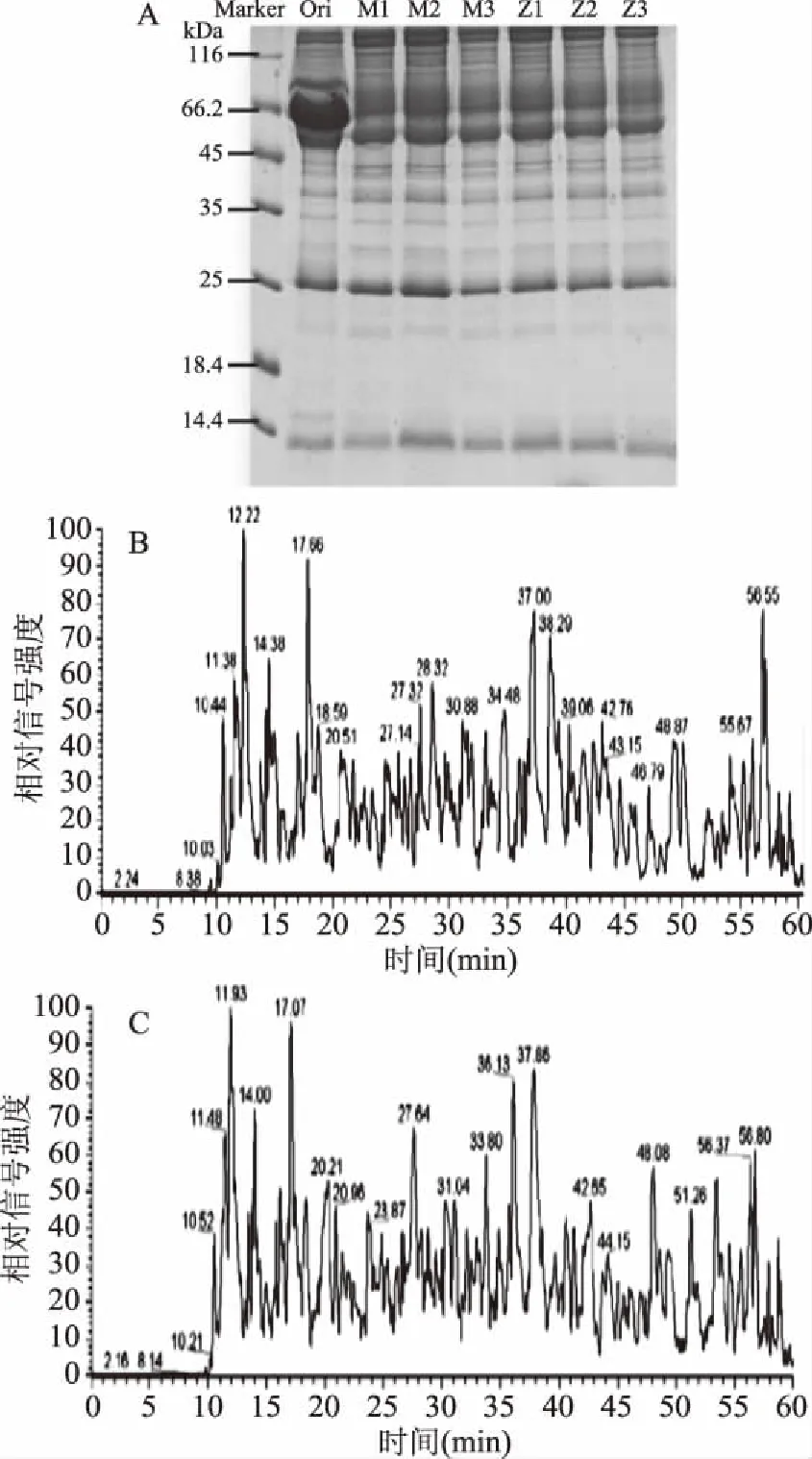
图1 小鼠血清样本平行性Fig.1 Samples parallelism of proteins in serum in mice注:A:小鼠血清蛋白SDS-PAGE; B:去除高丰度蛋白的模型组M1小鼠血清蛋白质谱图; C:去除高丰度蛋白的给药综合组Z1小鼠血清蛋白质谱图。
2.5 蛋白质鉴定结果统计
通过Maxquant中的Label free算法对二级质谱数据进行非标记定量计算结果如表3,3个模型组鉴定到肽段数分别为3101、3352、3328个,查库分别得到295、312、321种蛋白质;3个灌胃鲟鱼肽组鉴定到肽段数分别为3335、3299、3320个,查库分别得到321、308、317种蛋白质。

表3 蛋白质鉴定结果统计Table 3 Statistic of identified proteins
通过GO功能注释,结果如图2。小鼠低丰度蛋白参与了21个生理过程,其中蛋白质数目较多的是单生物过程、代谢过程、细胞过程;具有7种分子功能,主要是具有结合和催化活性功能;存在于10种细胞组分中,主要在细胞部分、细胞、细胞器和细胞膜外区中。

图2 基因本体2水平统计Fig.2 GO level 2 statistics
对参与的生物学过程、具有的分子功能和所处的细胞组分对小鼠血清低丰度表达蛋白质进行分类,通过Fisher精确检验评价小鼠血清低丰度表达蛋白质显著富集度(表4)。结果表明,在小鼠血清低丰度表达蛋白质中,与M组相比,Z组有4、2和2种显著表达的蛋白质分别参与了糖代谢过程、活性氧响应和细胞氧化还原3个生物过程,其富集度分别为0.19、0.4和0.33;而参与信息素结合、小分子结合、气味结合、转运、异构化、糖苷键水解和糖化合物水解等7个生物功能的蛋白质分别有3、5、3、4、2、2、2种表达显著。
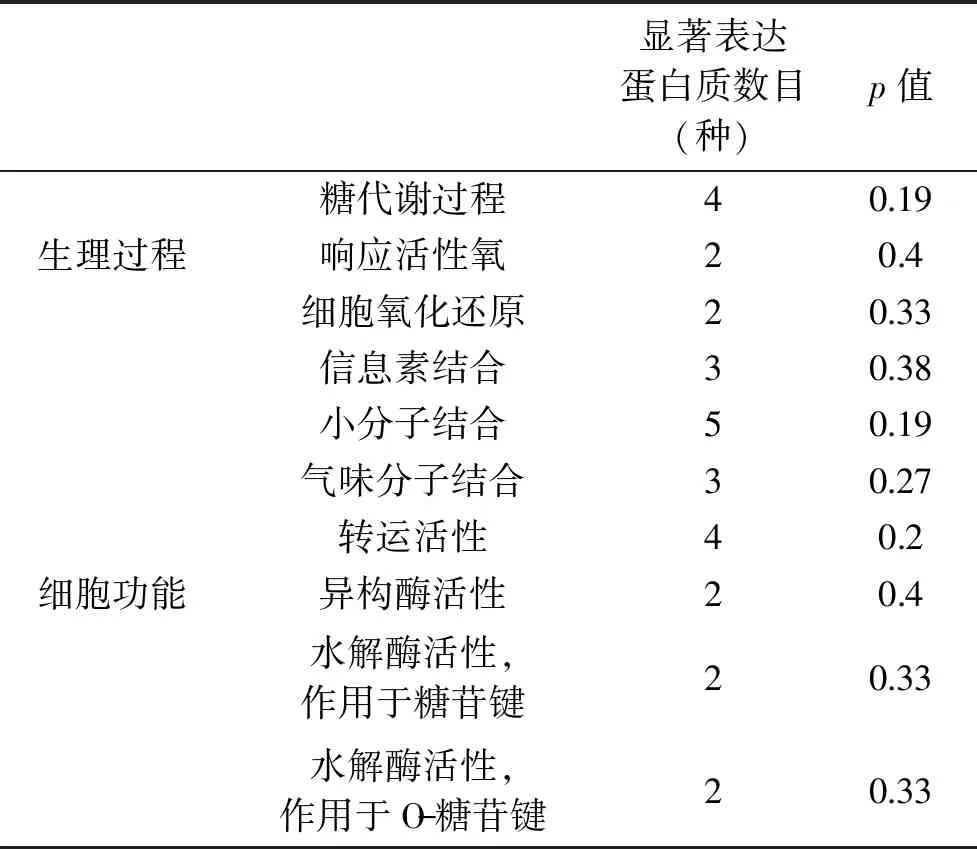
表4 显著富集的GO term统计Table 4 Enriched GO terms
在模型组的3个平行组和在灌胃鲟鱼肽的3个平行组中,筛选出同一平行组中至少有两个非空值的数据,同时根据蛋白质表达差异倍数>2.0倍(上调或下调)且p值<0.05的筛选标准,获得差异表达蛋白质3个,其中1个上调,2个下调(表5),即二磷酸甘油酸变位酶表达量显著上调(Z/M为2.07,p<0.05),间α胰蛋白酶抑制剂H4和主要尿蛋白20表达量显著下调(Z/M分别为0.38和0.22,p<0.05)。根据蛋白质表达差异倍数>1.5且p值<0.05的筛选标准,获得9个蛋白质(5个上调,4个下调)进行生物信息分析(表5),其中5种表达上调的蛋白质为二磷酸甘油变位酶、载脂蛋白D、黄素还原酶、纤蛋白-1和过氧化物酶-2,4种表达下调的蛋白质分别为巯基氧化酶1、主要尿蛋白1、间α胰蛋白酶抑制剂H4和主要尿蛋白20。这9种蛋白质中,二磷酸甘油酸变位酶、黄素还原酶和过氧化物酶-2分别参与糖酵解途径、卟啉和叶绿素代谢途径、细胞凋亡途径。二磷酸甘油酸变位酶、载脂蛋白D、黄素还原酶和过氧化物酶-2的上调以及巯基氧化酶1的下调可以提高机体清除自由基的能力。

表5 进行生物信息学分析的9种蛋白质Table 5 Statistic of the 9 proteins for bioinformatics analysis
生物体内的各种蛋白质在行使其生物功能时,常常需要其他相关蛋白质参与。将这些相互作用的蛋白质以网络形式连接,从而获得蛋白质交互作用网络。灌胃鲟鱼肽小鼠差异表达蛋白质的交互作用网络见图3。从图中可以看出,主要尿蛋白1(AC:Q4FZE8,基因名Mup1)、间α胰蛋白酶抑制剂H4(AC:A6X935,基因名Itih4)、二磷酸甘油酸变位酶(AC:P15327,基因名Bpgm)、黄素还原酶(AC:Q923D2,基因名Blvrb)、过氧化物酶-2(AC:Q61171,基因名Prdx2)、纤蛋白-1(AC:Q08879,基因名Fbln1)分别与4、4、1、2、9和1种蛋白具有交互作用。

图3 差异表达蛋白质相互作用网络Fig.3 PPI networks of the 9 proteins
3 结论
本研究通过D-半乳糖致小鼠衰老实验,发现鲟鱼肽可以引起小鼠体重、脾指数、血清中T-AOD和SOD活力增加,并同时降低MDA含量。去除高丰度蛋白后的小鼠血清样本中,鲟鱼肽可以导致小鼠血清中二磷酸甘油酸变位酶、载脂蛋白D、黄素还原酶和过氧化物酶-2的表达上调,并导致巯基氧化酶1表达量下调,这些蛋白质表达量的增加或降低,可以提高机体清除自由基的能力,延缓小鼠衰老过程。这些结果表明,分子量为805~3300的鲟鱼肽具有抗小鼠衰老作用。
猜你喜欢
古今农业(2021年2期)2021-08-14 01:43:14
中成药(2018年8期)2018-08-29 01:28:24
中成药(2018年3期)2018-05-07 13:34:11
大自然探索(2017年1期)2017-02-14 00:13:36
中国比较医学杂志(2017年3期)2017-01-17 05:40:54
中华老年多器官疾病杂志(2016年7期)2016-04-28 08:43:12
山东医药(2015年15期)2016-01-12 00:39:58
国际心血管病杂志(2015年5期)2015-02-27 12:11:33
山东畜牧兽医(2014年10期)2014-04-05 14:53:34
华东理工大学学报(自然科学版)(2014年4期)2014-02-27 13:49:12
