
滨海人工林土壤呼吸各组分对台风强降雨的响应
2019-03-25 05:28:10王磊桑昌鹏余再鹏林宇施秀珍万晓华黄志群
亚热带资源与环境学报 2019年1期
王磊,桑昌鹏,余再鹏,林宇,施秀珍,万晓华,黄志群*
(1.福建师范大学a.湿润亚热带山地生态国家重点实验室培育基地,b.地理科学学院,福州 350007;2.中国科学院 沈阳应用生态研究所,沈阳 110016;3.福建省长乐大鹤国有防护林场,福州 350212)
0 引言
陆地生态系统土壤呼吸每年向大气排放约68~98×1015g碳, 是化石燃料燃烧释放CO2的10~13倍[1-3],其微小波动都会对大气CO2浓度产生显著影响 ,进而直接影响全球碳平衡[4]。 IPCC第五次评估报告指出近百年来全球极端降水事件增加[5],而土壤的水分条件是影响土壤呼吸的主要环境因子之一,在降水改变的背景下,研究土壤水分改变对土壤呼吸的影响,有利于预测土壤碳排放对降水改变的响应规律。
土壤呼吸由异养呼吸(Rh)和自养呼吸(Ra)组成,Rh来源于微生物驱动下土壤有机质和植物残留物的分解,受底物的数量和质量调控[6-7];Ra来源于根系和根际微生物呼吸,与地上部分的光合作用密切相关[8-10]。研究表明,土壤呼吸各组分对降雨响应的时间存在很大差异:降雨迅速改变了地表凋落物的水分状况,湿润的环境使其分解速率加快,地表凋落物和土壤有机质分解(即异养呼吸)对降雨响应很快,大约需要1小时到数小时;而根呼吸(即自养呼吸)依赖于地上部分的光合作用,响应较慢,需要几天到几周[8]。有研究指出土壤自养呼吸与植物前三天的光合有效辐射显著相关[11],而且与土壤总呼吸、异养呼吸相比,自养呼吸对土壤湿度的响应不太敏感[12-13]。以往的研究表明,在土壤水分条件改变时,Ra/Rh的比例往往也会随之变化[14],刘博奇等[15]在小兴安岭典型温带森林土壤呼吸对强降雨响应的研究中也发现Ra、Rh对水分响应的不同步现象。因此,只有更深入地研究土壤呼吸各组分对降雨的响应规律,才能更好地理解土壤呼吸对降雨的响应过程,在更大尺度上预测土壤呼吸对降雨的响应趋势[16]。国内外关于土壤呼吸对降水响应的研究已取得一些进展[17-20],但这些研究都是通过控制实验模拟降雨对土壤呼吸的影响,不能完全反映自然状态下极端降水事件对土壤呼吸的影响。即使目前土壤呼吸对自然状态下强降雨响应的研究取得一些进展[21-22],但他们的研究多集中关注土壤总呼吸对降水改变的响应。Chen 等[11]通过区分土壤呼吸组分来研究玉米田自养呼吸和异养呼吸对极端降雨和降雪响应的规律,但是,在森林生态系统中,关于不同组分土壤呼吸对强降雨响应的研究仍鲜有报道[16]。
台风是影响亚热带滨海森林生态系统的一种常见的自然干扰,导致林内土壤温度[23]、水分[24]等环境因子在短时间内发生急剧变化,进而影响土壤呼吸速率。滨海地区多沙地人工林,沙地土壤的固相骨架松散、砂粒多而黏粒少、粒间孔隙大、降水容易渗入、内部排水迅速、蒸发失水强烈、水汽由大孔隙扩散到土表丢失,蓄水能力弱[25]。这些属性使滨海沙地具有不同于内陆森林生态系统的土壤环境。针对这些特殊条件,于2015年夏季在“苏迪罗”和“杜鹃”2次台风强降雨前后,分别对设置去除根系和对照处理一年后的湿地松(Pinuselliottii) 和尾巨桉(Eucalyptusurophylla×Eucalyptusgrandis)人工林监测土壤呼吸速率和相关环境因子,研究不同组分的土壤呼吸对台风强降雨的响应特征及规律,并探讨其相关机制,为理解和预测土壤呼吸对极端降雨的响应提供科学依据。
1 研究区自然概况
试验样地位于福建省长乐市大鹤国有防护林场(25°57′N,119°24′E),地处福建省东部沿海、闽江口南岸,东濒台湾海峡,属于沿海防护林基干林带区域。本试验样地气候属典型的亚热带海洋性季风气候,雨量充沛,气候温和,多年平均气温19 ℃,最高气温35 ℃,最低气温为0 ℃,多年平均降水量1 383 mm。年实际日照时数1 837 h,无霜期326 d,空气相对湿度达80%。平均海拔为10 m,土壤为滨海风积沙土,沙层深厚,可达6~10 m。土壤肥力较差,保水能力较低。选取湿地松和尾巨桉2个树种为研究对象,分别作为针叶、阔叶树种的典型代表。湿地松造林于1991年,尾巨桉是在湿地松采伐迹地上于2003年营造的。林下植被主要有牛口刺(Cirsiumshansiense)、白茅(Imperatacylindrica)、鸭跖草(Commelinacommunis)和天门冬(Asparagussprengeri)。2个人工林样地基本概况和0~10cm土层内的细根及土壤性质见表1。
2 研究方法
2.1 样地设计

表 1 湿地松和尾巨桉人工林林地基本概况
注:同一行不同小写字母表示2个树种之间差异显著(P<0.05)。
2014年9月,在2个树种人工林内分别设置4个20 m×20 m的小区,然后按照随机区组设计方法,在每个小区内分别设置2个2 m×2 m的次小区,作为2个处理:1)去除根系(Root-trenching,RT),沿着小区四周挖1 m深的壕沟断根进行组分分离,并在壕沟内埋入 3层孔径为0.1 mm的尼龙网阻断周围根系向小区内生长,并在监测期间去除此小区内活体植物;2)对照(Control,CK),即不作任何处理。在各次小区内埋设土壤呼吸圈,呼吸圈为直径20 cm,高10 cm 的PVC管,楔入土壤后地上部分保留2 cm,整个研究期间呼吸圈位置保持不变。
2.2 土壤呼吸速率及温、湿度的测定
利用开路式土壤碳通量测定系统(Li-8100,美国)监测各次小区内土壤呼吸速率,设定每个呼吸圈的监测时间为2 min;采用便携式数字温度计测定土壤呼吸圈附近地下10 cm 处的土壤温度;采用时域反射仪(TDR)测定0~10 cm 范围内土壤的体积含水量,土壤温、湿度在每个土壤呼吸圈附近测定3次取平均值。土壤呼吸速率与土壤温、湿度同时监测,观测时间选择在2次台风强降雨发生前后每天上午的9:00—11:00监测一次。本研究的2次数据来源于2015年8—10月份2次台风强降雨发生前后的测量,日期分别是8月4日和8月9日—13日和9月28日和9月30日—10月3日,2次台风强降雨过程中未进行监测。
2.3 台风强降雨
样地地处福建省东部沿海,每年多发台风,分别选取2015年8月、9月各一次台风强降雨前后进行研究。2015年第13号台风“苏迪罗”于8月7日开始影响中国东南沿海,受其影响,福建省长乐市8月7、8、9日3天的降水量分别为19.8、261.6和46.5 mm。2015年第21号台风“杜鹃”于9月28日上午开始影响中国东南沿海,受其影响,福建省长乐市9月28、29、30日及10月1日4天的降水量分别为52.3、114.1、5.6和7.1 mm。9月28日土壤呼吸及土壤温、湿度的监测在台风强降雨之前。为方便表述,2次台风强降雨依次简称为第一次、第二次降雨,具体情况如下(表2)。
2.4 数据统计分析

表 2 2次台风强降雨的基本情况
土壤呼吸不同组分的计算:
Ra=Rs-Rh
(1)
式(1)中:Ra为自养呼吸(μmol CO2·m-2·s-1),Rs为对照小区土壤呼吸速率即总呼吸,Rh为去除根系小区土壤呼吸速率即异养呼吸。
异养呼吸的土壤湿度W1(%)均为去根小区土壤的湿度,自养呼吸的土壤湿度W2(%)均为对照小区土壤的湿度,土壤呼吸速率与土壤湿度(W) 之间的关系通过二次多项式(2)拟合[22]:
R=aW2+bW+c
(2)
式(2)中:R为测定的土壤呼吸速率,W为地下0~10 cm土壤湿度,其他字母均为待定参数。
采用Excel 2010和SPSS 19.0 软件进行数据处理和统计分析,由Origin 9.0软件作图描述降雨前后的土壤呼吸速率的变化,用二次多项式关系模型来拟合台风强降雨前后土壤呼吸各组分和0~10 cm土壤湿度之间的单变量关系。
3 结果与分析
3.1 台风强降雨前后土壤呼吸各组分及土壤湿度的变化
图1是湿地松和尾巨桉人工林土壤总呼吸、异养呼吸和自养呼吸对2次台风强降雨的响应及土壤湿度的变化。2次台风降雨前,样地在副热带高压控制下持续干旱,第一次降雨分别使湿地松和尾巨桉对照小区土壤湿度比降雨前提高了93%和83%,第二次降雨分别使其比降雨前提高了16%和18%,降雨显著改善了土壤水分状况。第二次降雨前湿地松和尾巨桉对照小区土壤湿度分别为4.5%和5.1%,显著高于第一次降雨前湿地松和尾巨桉对照小区土壤湿度(P<0.01)。第一次降雨最后一天,湿地松和尾巨桉2种人工林对照和去根小区的土壤湿度仍未恢复到降雨前水平,而第二次降雨结束后土壤湿度迅速恢复。

图 1 2次台风强降雨前后土壤呼吸各组分及土壤湿度的变化Figure 1 Soil respiration components and soil moisture changed with time around the two typhoons with great rainfall

图 2 土壤自养呼吸、异养呼吸速率占总呼吸的比例Figure 2 The contribution of autotrophic respiration and heterotrophic respiration rate to soil respiration
2次台风降雨后,湿地松人工林土壤总呼吸、异养呼吸速率均呈现出快速上升,然后逐渐下降的趋势,而土壤自养呼吸速率的上升则不明显。第一次降雨后,湿地松土壤总呼吸、异养呼吸和自养呼吸速率分别比降雨前提高70%、90%和30%,尤其是土壤异养呼吸速率增加了1.44 μmol CO2·m-2·s-1。第二次降雨后,湿地松土壤总呼吸、异养呼吸和自养呼吸速率分别比降雨前提高40%、30%和60%,土壤异养呼吸速率在降雨开始后第3天迅速达到峰值。土壤总呼吸和异养呼吸的峰值与土壤湿度的峰值同步,但自养呼吸的峰值比土壤湿度峰值要滞后1天。
尾巨桉人工林在2次台风降雨后土壤总呼吸、异养呼吸速率也同样表现出快速上升、然后逐渐下降的趋势,而土壤自养呼吸速率的上升则不明显。第一次降雨后,其土壤总呼吸、异养呼吸和自养呼吸速率分别比降雨前提高140%、160%和70%,尤其是土壤异养呼吸速率增加了3.82 μmol CO2·m-2·s-1。在第二次降雨后,尾巨桉人工林土壤总呼吸、异养呼吸和自养呼吸速率分别比降雨前提高20%、20%和10%,尤其是土壤异养呼吸速率增加了0.92 μmol CO2·m-2·s-1。2次降雨尾巨桉土壤总呼吸及异养呼吸速率都在降雨结束1天后达到峰值,与土壤湿度峰值同步,而自养呼吸速率峰值在降雨结束2天后才达到峰值。
降雨前的持续干旱使得土壤水分处于亏缺状态,土壤呼吸各组分均受到了水分不足的制约。2次台风降雨均显著地促进了湿地松和尾巨桉土壤呼吸各组分,第一次降雨结束5天后湿地松和尾巨桉土壤呼吸各组分仍未恢复到降雨前水平,且2个树种土壤总呼吸、异养呼吸速率与降雨前差异显著(P<0.05)。而在第2次降雨结束后的第1天就迅速恢复甚至低于降雨前水平。尾巨桉土壤呼吸各组分在2次台风强降雨中均显著高于湿地松(P<0.05)。
3.2 土壤自养呼吸、异养呼吸对台风强降雨的响应
图2是强降雨前后土壤自养呼吸、异养呼吸速率占总呼吸的比例随时间的变化。2次台风强降雨导致湿地松和尾巨桉人工林土壤异养呼吸速率快速增加,显著提高了异养呼吸/总呼吸,第一次降雨使湿地松土壤异养呼吸/总呼吸由66.86%(降雨前8月4日)提高到83.73%(降雨结束第2天),土壤自养呼吸/总呼吸的变化范围为16.27%~40.19%。尾巨桉土壤异养呼吸/总呼吸由75.33%(降雨前1天)提高到86.27%(降雨结束当天),土壤自养呼吸/总呼吸的变化范围为13.73%~24.67%。观测期间,湿地松和尾巨桉土壤异养呼吸对总呼吸的贡献率明显大于自养呼吸,其贡献率分别是自养呼吸贡献率的5.15倍和6.28倍。2个人工林中,湿地松土壤自养呼吸对总呼吸的贡献率显著高于尾巨桉,而尾巨桉土壤异养呼吸对总呼吸的贡献率显著高于湿地松(P<0.05)。第二次降雨使得湿地松和尾巨桉人工林土壤异养呼吸/总呼吸也表现出与第一次相同的趋势。
3.3 台风强降雨前后土壤呼吸各组分与土壤湿度之间的关系
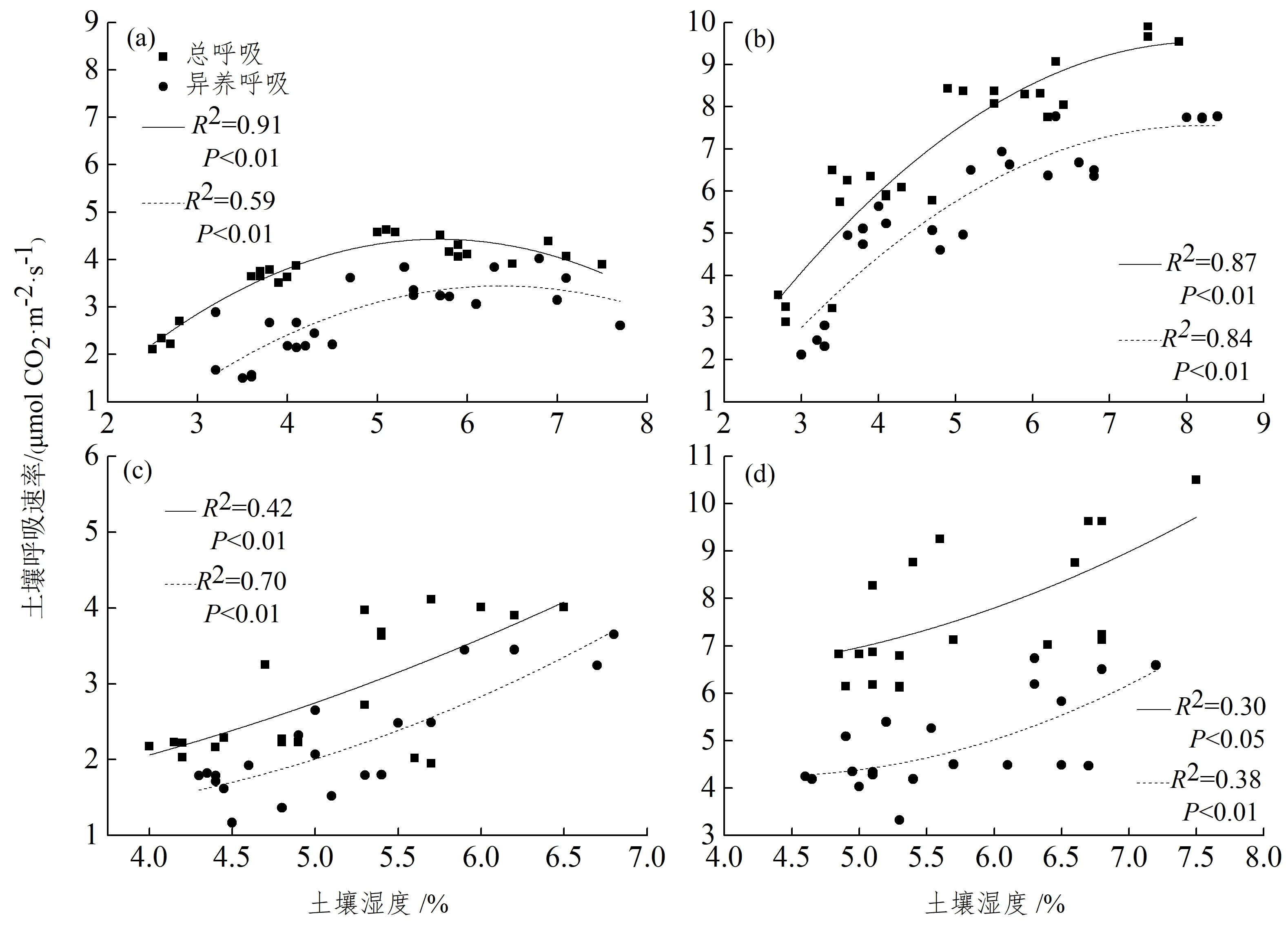
注:第一次降雨的湿地松(a)和尾巨桉(b),第二次降雨的湿地松(c)和尾巨桉(d)。图 3 2次台风强降雨前后,土壤呼吸速率和土壤湿度之间的关系Figure 3 The relationships between the soil respiration rate and soil moisture before and after great rainfall of two typhoons
图3反映了2次台风强降雨前后,土壤总呼吸及异养呼吸速率对土壤湿度的响应特征,用二次多项式拟合二者之间的关系。湿地松和尾巨桉人工林土壤总呼吸及异养呼吸在2次台风强降雨事件中与0~10 cm土壤湿度均显著相关。第一次台风强降雨前后6天范围内,湿地松人工林土壤总呼吸约63%的数值位于拐点值(5.69%)的左侧,其异养呼吸约83%的数值位于拐点值(6.36%)的左侧;而尾巨桉人工林土壤总呼吸全部位于拐点值(8.23%)的左侧,其异养呼吸约88%的数值位于拐点值(8.17%)的左侧;第二次台风强降雨前后5天范围内,湿地松和尾巨桉人工林土壤总呼吸及异养呼吸与土壤湿度之间的二次曲线均不存在拐点。2次台风强降雨中,第一次降雨湿地松土壤湿度对总呼吸、异养呼吸的解释范围分别为91%和59%,尾巨桉土壤湿度对总呼吸、异养呼吸的解释范围分别为87%和84%。第二次降雨湿地松土壤湿度对总呼吸、异养呼吸的解释范围分别为42%和70%,尾巨桉土壤湿度对总呼吸、异养呼吸的解释范围分别为30%和38%。第一次降雨土壤湿度对土壤总呼吸及异养呼吸的解释除湿地松异养呼吸外,明显高于第二次降雨。2次台风强降雨前后湿地松和尾巨桉人工林土壤自养呼吸和土壤湿度均无显著相关性(P>0.05)。
4 讨论
4.1 台风强降雨对土壤呼吸各组分的影响
土壤水分波动通常会引起生态系统物质循环的变化,这在干旱、半干旱与季节性干旱地区尤为明显[26-27]。土壤水分过多或不足均会抑制土壤呼吸,只有在土壤水分含量适中时土壤呼吸速率最大[28-29]。降雨对土壤呼吸的影响与土壤的初始水分状况密切相关,通常降雨能激发干燥土壤的呼吸,而抑制潮湿土壤的呼吸[16]。
研究在亚热带滨海沙地的湿地松和尾巨桉人工防护林,对发生在2015年的2次台风强降雨前后土壤呼吸速率及环境因子进行测定。结果表明,2次台风强降雨使得湿地松和尾巨桉人工林土壤总呼吸、异养呼吸和自养呼吸速率明显提高,这与Yu等的研究结果相似[30]。土壤呼吸各组分对降雨的响应存在很大差异[11]。研究中湿地松和尾巨桉人工林土壤异养呼吸和总呼吸对台风强降雨响应迅速且与土壤湿度同步,但自养呼吸对台风强降雨的响应要明显滞后于土壤总呼吸和异养呼吸。相关研究表明,土壤异养呼吸对环境因子变化的响应非常敏感,生物或非生物因子的微小变化将直接影响呼吸速率的大小[31],这可能是由于降雨迅速改变了地表凋落物的水分状况,使大量易变性有机底物淋溶释放出来,促进微生物分解;另一方面,可能与土壤有机质分解速率的变化有关,降水显著地改善了土壤湿度,使得微生物生物量增加,活性增强,促进了易分解有机底物的可利用性和扩散[18,32-33],导致CO2排放迅速增加。土壤异养呼吸对土壤湿度变化的响应比自养呼吸更为迅速,因为激发植物根系活性比激发表层土壤微生物群落需要更多的水分[34]。在干旱和半干旱地区,降雨可以引发植物的萌发与生长[27],沙漠植物在土壤加湿后需几天的时间才开始新根的生长[35],而干旱的土壤在雨后2个星期,植物叶片才变得更绿[36]。所以,降雨引起的土壤自养呼吸峰值滞后,对降雨的响应要慢于异养呼吸,土壤呼吸各组分对降水变化的敏感性不同,异养呼吸比自养呼吸更敏感。对于干旱或水分相对亏缺的土壤,降雨能够迅速激发土壤呼吸,通常这种激发效应可以持续2~6 天[37-38]。第一次台风强降雨结束4天后,湿地松和尾巨桉人工林土壤呼吸各组分仍未恢复到降雨前水平,这可能和第一次台风降雨量大(327.7 mm),且降雨前样地土壤长时间的持续干旱,湿地松和尾巨桉土壤总呼吸其湿度平均值分别仅有2.7%和2.9%,异养呼吸其湿度平均值分别仅有3.5%和3.2%有重要关系。但第二次台风强降雨结束2天后,湿地松和尾巨桉人工林土壤呼吸各组分就恢复到降雨前水平,这可能由于第二次降雨前土壤湿度高,雨量小(179.1 mm),且本研究区域为滨海沙地,相比于其他土壤质地,沙地土壤固相骨架松散,砂粒多而黏粒少,粒间孔隙大降水容易渗入,内部排水迅速,蒸发失水强烈,排水透气状况好,土壤气体扩散性强,易受水分胁迫[25],沙地的土壤呼吸恢复到降雨前的水平需要的时间短。本研究发现尾巨桉人工林土壤呼吸各组分显著高于湿地松,Janssens等指出不同森林类型间植被生产力的差异是影响土壤呼吸差异的最重要因素[39]。杨玉盛等也发现阔叶林土壤呼吸速率显著高于相同立地条件下的针叶林[40]。尾巨桉人工林土壤异养呼吸及自养呼吸高于湿地松,与湿地松相比,尾巨桉凋落物及细根质量高(C/N低,纤维素、木质素含量低),可能易分解淋溶转化成可溶性有机碳并提高土壤微生物生物量和活性,因而会刺激异养呼吸速率升高[41]。从土壤自养呼吸及异养呼吸占土壤总呼吸的比例来看,自养呼吸及异养呼吸对水分响应不同步,这与刘博奇等研究结果一致[16]。强降雨对土壤呼吸的促进作用主要表现在异养呼吸方面且响应迅速,而自养呼吸对降雨的响应要滞后于异养呼吸。湿地松和尾巨桉土壤异养呼吸、自养呼吸对总呼吸贡献率不同,这就意味着根系对土壤呼吸的影响因树种而异[42]。已有研究表明树种能调控地下碳分配的比例、根系周转速率及生物量而影响土壤呼吸速率[40]。
4.2 台风强降雨前后影响土壤呼吸各组分的关键环境因子
利用二次多项式拟合土壤呼吸速率与0~10 cm土壤湿度之间的关系(图3),以期阐释发生在2次台风强降雨前后影响土壤呼吸的关键因子。结果表明,第一次台风强降雨从整体上促进了湿地松人工林土壤总呼吸、异养呼吸,但其与土壤湿度的二次曲线出现了拐点,此拐点对应的土壤湿度可以作为判定促进或抑制作用的临界值[21]。当降雨对土壤水分的补给使土壤湿度小于临界值时,降雨会促进土壤总呼吸及异养呼吸;当超过临界值时,土壤总呼吸及异养呼吸就会降低,降雨就会对土壤呼吸产生抑制作用。相关研究表明:降雨会对林冠层和地表凋落物淋溶[43],促进根系生长和周转[16],产生大量简单且易分解的碳水化合物,共同提高微生物生物量和活性来促进土壤呼吸[44]。而降雨使土壤湿度超过土壤临界点时,土壤孔隙被填充,限制了CO2的扩散,厌氧条件降低了微生物生物量、活性和抑制根系及根际微生物活动等来降低土壤呼吸[11]。第一次台风强降雨整体上促进了尾巨桉人工林土壤总呼吸,却短暂的抑制了异养呼吸。桑昌鹏等在样地研究发现去除根系显著增加了各树种0~10 cm土壤含水量[45],这可能是由于去除根系的小区没有根系吸收水分导致土壤湿度大于对照小区且超过临界值(8.17%),所以才造成降雨对异养呼吸的短暂抑制。第二次台风强降雨对湿地松和尾巨桉人工林土壤总呼吸、异养呼吸均没有抑制,这可能是由于第二次台风强降雨累积降雨量(179.1 mm)比第一次(327.7 mm)少,且第二次台风强降雨的降雨强度要小于第一次。当然,由于技术原因不能够对降雨前后以及过程中的土壤呼吸变化进行全天候连续观测,可能会错过降雨过程中由于降雨过多导致的土壤呼吸抑制现象。2次台风强降雨前后湿地松和尾巨桉人工林的土壤自养呼吸均和土壤湿度无显著相关性(P>0.05)。这与Chen 等的研究结果相一致,其研究指出土壤自养呼吸与植物光合有效辐射显著相关,且年自养呼吸通量与地上生物量线性相关[11]。还有研究指出,与土壤总呼吸、异养呼吸相比,自养呼吸对土壤湿度的变化不敏感[12-13]。
5 结论
研究通过监测2次台风强降雨前后湿地松和尾巨桉滨海人工林土壤呼吸各组分和环境因子发现:土壤的初始水分状况、降雨量、土壤物理性质和树种都会影响到土壤呼吸各组分对降水改变的响应。湿地松和尾巨桉人工林土壤异养呼吸和总呼吸对台风强降雨响应迅速且与土壤湿度同步,但自养呼吸对台风强降雨的响应要明显滞后。当土壤湿度小于临界值时,降雨会促进土壤总呼吸及异养呼吸,当超过临界值时,土壤总呼吸及异养呼吸就会降低,但土壤湿度不是影响自养呼吸的关键因子。这些发现对土壤碳通量及其对气候变化的反馈具有重要意义,因为随着气候变暖,极端天气(干旱、暴雨和台风等)在全球范围内将更加频繁和强烈。因此,极端降雨对土壤碳循环的影响应纳入到地球系统模型以提高对C-气候反馈的预测。
目前,土壤呼吸对降雨响应的研究还不够深入,需要进一步区分土壤呼吸各组分,在较大时空尺度上定量研究呼吸对降雨的响应,考虑其他全球变化因子,更多地关注土壤微生物和植物生理方面(如光合作用)。
猜你喜欢
农业与技术(2021年6期)2021-12-12 05:33:47
天津农林科技(2020年3期)2020-08-13 03:17:10
生物工程学报(2020年6期)2020-07-31 08:48:32
林业与生态(2020年2期)2020-04-16 12:48:31
电子制作(2019年15期)2019-08-27 01:12:12
广西林业科学(2016年3期)2016-03-16 05:43:30
高原山地气象研究(2016年4期)2016-02-28 13:53:39
高原山地气象研究(2016年4期)2016-02-28 13:53:37
生物灾害科学(2015年3期)2015-03-11 16:38:36
农家科技下旬刊(2014年11期)2015-02-09 19:53:12
