
毛乌素沙地植被类型对生物结皮及其下伏土壤养分的影响
2019-03-25 07:55董金伟李宜坪李新凯李炳垠卜崇峰2
水土保持研究 2019年2期
董金伟, 李宜坪, 李新凯, 李炳垠, 郭 琦, 卜崇峰2,
(1.北京中邦文旅科技发展有限公司, 北京 100083; 2.中国科学院 水利部水土保持研究所, 陕西 杨凌 712100; 3.中国科学院大学, 北京 100049; 4.西北农林科技大学水土保持研究所, 陕西 杨凌 712100; 5.西北农林科技大学 水利与建筑工程学院, 陕西 杨凌 712100)
生物结皮对极度干旱、营养物质匮乏的环境条件有很强的适应能力,能通过光合作用、生物固氮增加土壤养分[1-3],并通过减少土壤侵蚀而减轻土壤中的养分流失。生物结皮的养分积累作用主要表现在结皮层及其下伏0—5 cm土层,对深层土壤的有机质、全氮及速效养分作用并不大[4-5]。生物结皮的覆盖对植物的生长发育以及生态系统的演变具有重要的影响,能够为维管束植物的生长和定居提供有利的生存条件[6-8,10]。反过来,维管植物能够沉积足够的黏粒和粉粒,并为结皮的生长提供适宜的水分环境[9]。同时,植物可以减少干扰并提供遮荫条件,促进生物结皮的生长[3]。已有研究多为植被与生物结皮的共生关系探讨[9-10],而有关植被类型及其空间分布对生物结皮土壤养分的影响认识还十分缺乏,针对毛乌素沙地这一独特的地理单元的相关研究也鲜有报道[5,11-12]。为此,本文选择毛乌素沙地苔藓及藻结皮发育较好的4种典型植被样地,探讨植被冠层对生物结皮及其下伏土壤养分特征的影响,为沙漠地区生态建设及荒漠化防治提供借鉴。
1 材料与方法
1.1 研究区概况
本研究选择毛乌素沙地样地12个,散布在内蒙古自治区乌审旗南部及鄂托克前旗东北部(图1)。毛乌素沙地(37°20′—39°23′N,107°20′—111°30′E)处于鄂尔多斯高原南部向陕北黄土高原的过渡地区。地势自西北向东南倾低,平均海拔1 300~1 600 m。地形以丘陵、梁地、洪积—冲积台地、谷地和滩地为主。沙区大部分属于温带大陆性半干旱气候,年均温6.0~8.5℃。年均降水量东南部为440 mm,向西递减至250 mm,降雨主要集中在7—9月,占全年总降水量的60%~75%。年蒸发量为1 800~2 500 mm。年均日照时数西北部为3 000~3 100 h,东南部降至2 800~2 900 h,太阳辐射总量从西北部的627.6 kJ/(cm2·a)逐渐降至东南部的577.6 kJ/(cm2·a)。≥10℃积温2 700~3 000℃。沙区盛行风向为西北风,年平均风速4.8 m/s,年平均大风日数10~40 d。气候干燥、冷热剧变、大风频繁、日照强烈是该地区的主要气候特征。
毛乌素沙地沙生植被类型主要有白沙蒿(Artemisiasphaerocephala)、黑沙蒿(Artemisiaordosica)、杨柴(Hedysarumlaeve)、柠条(Caraganaintermedia)、沙地柏(Sabinavulgaris)、沙柳(Salixpsammophila)、乌柳(Salixmicrostachya),除以上集中沙生植物外在沙区东南隅还有连片的灌木丛,如小叶鼠李(Rhamnusparvifolius)、丝绵木(Euonymusbungeanus)等。
图1典型植被群落土壤及生物结皮采集地
1.2 研究方法
基于全面野外调查,选取沙蒿、沙柳、柠条、草地4类典型植被群落。每种类型植被设3个重复,样地大小为30 m×30 m,样地实景如图2所示,样地基本概况见表1。
试验设计见表2,根据生物结皮类型,在每个样地内设置:主导植被冠层下方苔藓及藻结皮,植被冠层间苔藓及藻结皮,并设置裸地对照。苔藓结皮用环刀获取结皮层,并用微型土钻分层取结皮层下伏0—5 cm,5—10 cm土壤;藻结皮自地表以下分层取0—5 cm,5—10 cm土壤。均为3个重复,采用五点采样法取混合样,装入自封袋带回室内,土样均过0.01 mm筛。土壤有机质、全氮、全磷含量均采用全自动间断化学分析仪测定(CleverChem 200),每个样品均测定3个重复值。
1.3 数据处理与分析
试验数据采用样本均值加减3倍标准差识别特异值,在此区间外的数据均定为特异值,分别用正常的最大值和最小值代替,后续计算均采用处理后的原始数据。用Microsoft Excel 2010做整理后,采用SPSS 19.0统计软件及SigmaPlot 12.5绘图软件对试验数据进行方差分析和绘图。

图2 研究区各群落样地实景图

植被类型海拔高程/m坡度/(°)优势建群种植被盖度/%藻结皮盖度/%藓结皮盖度/%沙蒿13184黑沙蒿、沙柳、花棒692312沙柳11906沙柳、黑沙蒿、骆驼蓬1547柠条12641黄蒿、柠条3532草地12800草木樨、针茅草5585

表2 试验设计
2 结果与分析
2.1 不同植被群落生物结皮及其下伏土层的有机质分布
生物结皮可以显著增加土壤表层有机质等土壤养分的含量,且不同的植被群落中发育的生物结皮,其下伏土壤有机质等含量差异也很大[13-14]。通过对沙柳、沙蒿、柠条群落苔藓及藻结皮的有机质含量分布分析(图3),可知研究区典型植被群落对生物结皮及其下伏土壤的养分累积有显著影响。其中,3类植被群落下的苔藓结皮层有机质含量显著高于下伏土层,包括草地在内的各植被群落藻结皮下伏土壤亦表现出随土层加深有机质含量下降的趋势。生物结皮的发育位置对土壤养分的累积效应也不同。沙柳冠层间苔藓结皮及藻结皮下伏土层有机质含量均显著高于冠层下(p<0.05);沙蒿冠层下苔藓结皮层及下伏土层有机质含量也低于冠层间(p<0.05),藻结皮则相反;柠条冠层下苔藓结皮层有机质含量显著高于冠层间(p<0.05),而5—10 cm土层苔藓及藻结皮均为冠层下低于冠层间;较其他3种植被群落,草地苔藓结皮层有机质最低,且随着土层的加深,其有机质含量总体呈现增加的趋势,5—10 cm土层平均为(17.35±0.31) g/kg远高于苔藓结皮层有机质(3.97±0.10) g/kg,较之0—5 cm土层有机质增加了3.97倍,且藻结皮0—5 cm,5—10 cm土层有机质含量均显著高于苔藓结皮(p<0.05)。
总体上,有机质含量表现为:苔藓结皮>藻结皮>裸土,这与生物结皮的演替规律相一致。4种植被样地苔藓结皮有机质含量分别为:柠条(27.67 g/kg)>草地(24.81 g/kg)>沙柳(22.43 g/kg)>沙蒿(21.22 g/kg);藻结皮有机质含量则为:草地(13.61 g/kg)>柠条(12.37 g/kg)>沙蒿(9.60 g/kg)>沙柳(5.95 g/kg)。
2.2 不同植被群落生物结皮及其下伏土层的全氮分布
图4表明,沙柳、沙蒿、柠条3种植被群落下伏土壤的全氮含量均表现为随土层的加深而下降。从生物结皮发育位置来看,沙柳群落藻结皮和苔藓结皮下伏土壤全氮均为冠层间高于冠层下,冠层间苔藓结皮层全氮为1.02 g/kg,冠层下仅为0.45 g/kg;冠层下苔藓和藻结皮的同层下伏土壤全氮也有显著差异,0—5 cm土壤层苔藓结皮全氮平均为0.15 g/kg,显著高于藻结皮同土层全氮(0.13 g/kg),而5—10 cm土壤层苔藓和藻结皮全氮含量相当。沙蒿群落苔藓结皮表现为冠层间全氮高于冠层下,藻结皮与之相反;垂直方向上,冠层间苔藓结皮层及下伏土壤全氮均显著高于冠层下同层土壤(p<0.05),藻结皮0—5 cm,5—10 cm土层全氮则表现为冠层下高于冠层间;对比冠层下苔藓及藻同层土壤全氮发现:藻结皮下伏土壤全氮均显著高于苔藓结皮(p<0.05),冠层间0—5 cm土壤亦表现如此,而5—10 cm土壤藻结皮远低于苔藓结皮。柠条灌丛苔藓与藻土壤全氮变化表现不一:苔藓结皮层全氮为冠层下高于冠层间,0—5 cm土层全氮冠层下(0.34 g/kg)略高于冠层间(0.32 g/kg),而5—10 cm土层全氮冠层下(0.22 g/kg)低于冠层间(0.35 g/kg);藻结皮下伏0—5 cm,5—10 cm全氮变化特征与苔藓结皮一致。草地苔藓结皮表现出不同的特征,随土层加深,苔藓结皮下伏5—10 cm层土壤全氮(0.93 g/kg)远高于结皮层(0.20 g/kg)及0—5 cm层土壤(0.18 g/kg),而藻结皮下伏土壤随土层加深全氮逐渐降低。

图3各植被样地生物结皮及下伏土壤有机质分布
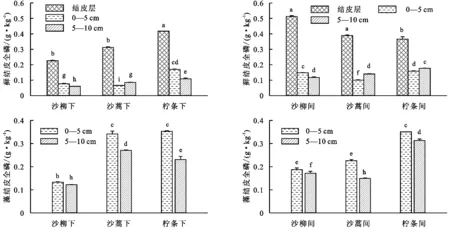
图4各植被样地生物结皮及下伏土壤全氮分布
总体上,全氮含量表现为:苔藓结皮>藻结皮>裸地,4种植被群落苔藓结皮土壤全氮为:柠条(1.40 g/kg)>草地(1.31 g/kg)>沙柳(1.14 g/kg)>沙蒿(1.09 g/kg);藻结皮土壤全氮为:草地(0.72 g/kg)>柠条(0.62 g/kg)>沙蒿(0.49 g/kg)>沙柳(0.31 g/kg);裸地对照全氮为:草地(0.59 g/kg)>柠条(0.41 g/kg)>沙蒿(0.30 g/kg)>沙柳(0.13 g/kg),与藻结皮表现出同样的特征。
2.3 不同植被群落生物结皮及其下伏土壤的全磷分布
杨永胜等[15]通过研究发现,沙柳及沙蒿植被苔藓结皮均能够显著提高土壤全磷含量,且主要集中在表层土壤,改善程度同植被类型具有密切的关系。由本次研究中各植被群落苔藓及藻结皮全磷分布(图5)可知,沙柳群落生物结皮发育位置不同其全磷变化表现不一。冠层间苔藓、藻结皮及下伏土层全磷均显著高于冠层下,冠层下苔藓结皮随土层加深,其全磷含量逐渐增加,而冠层间表现出相反趋势。沙蒿群落苔藓和藻结皮均表现出随土层加深,其全磷含量逐渐降低;冠层间苔藓结皮层及下伏土层全磷高于冠层下,藻结皮各土层全磷含量与冠层下相当。柠条群落冠层下与冠层间藻结皮各土层全磷含量差异不显著;苔藓结皮全磷表现出冠层下高于冠层间,冠层下随土层加深土壤全磷呈现降低的态势,而冠层间则表现为逐渐增加。草地苔藓、藻结皮土壤全磷均表现为上层土壤高于下层土壤,其中苔藓5—10 cm土层全磷累积量平均为(0.90±0.01) g/kg,高出0—5 cm土层磷素含量3.1倍。

图5各植被样地生物结皮及下伏土层全磷分布
总体上,全磷含量表现为苔藓结皮>藻结皮>裸地对照。就苔藓结皮来说,其土壤全磷为草地(1.35 g/kg)>柠条(1.02 g/kg)>沙蒿(0.63 g/kg)>沙柳(0.49 g/kg),藻结皮土壤全磷为草地(0.55 g/kg)>柠条(0.51 g/kg)>沙蒿(0.40 g/kg)>沙柳(0.28 g/kg),裸地对照则为草地(0.54 g/kg)>柠条(0.37 g/kg)>沙蒿(0.29 g/kg)>沙柳(0.22 g/kg)。
2.4 讨 论
上述结果分析显示,就生物结皮相对于植被灌丛的发育部位来说,冠层间生物结皮积累的土壤养分明显高于冠层下方,表明植被灌丛虽可为生物结皮提供较好的水分及遮荫条件,但生物结皮中苔藓等植物的存在势必与植被灌丛形成竞争关系,且植被灌丛的生长必然会消耗生物结皮积淀的碳、氮、磷等元素,而冠层间空地均在植被灌丛的根系半径之外,结皮下方的土壤养分不会被植灌根部所汲取,再者沙地结皮首先在冠层下方或冠层附近形成,然后才扩展至植物行间或空地[6],故冠层下大多为发育初期或中期的生物结皮,而冠层间则以发育稳定期的结皮居多,其结皮层及下伏土壤的土壤养分积累更多。
已有研究表明,不同植被群落中土壤养分的垂直分布规律差异较大[16]。本研究中,除苔藓结皮的全氮、有机质外,各植被群落中生物结皮及下伏土壤养分总体上表现为草地最高,柠条、沙蒿次之,沙柳林最低。原因是草地土壤通常容重较大,沙土含量较少,且雨季多有积水,水分条件较好,生物结皮虽覆盖度不高,但在草地土壤发育较为稳定,草本植物的生长不会消耗过多表层土壤养分,且不易受到破坏,结皮下土壤养分可以较好地累积[17];柠条样地土壤容重同样较大,土壤中砂粒含量较少,使生物结皮可以较稳定的发育,且柠条的根系较发达,其固氮能力较强,能够对土壤起到一定的改良效果[18];沙蒿群落土壤容重较小,砂粒含量居多,植被间受干扰踩踏较严重,导致生物结皮养分不易累积[19];沙柳林样地植被盖度均较低,冠层间少有生物结皮发育,且土壤砂粒较多,不利于结皮层的稳定发育[20],故土壤养分含量均较少。
3 结 论
本研究表明,植被类型与冠层覆盖对生物结皮及其下伏土壤层的养分含量及剖面分布特征有着显著的影响。表现在沙柳及柠条群落冠层间苔藓、藻结皮层及其下伏土壤的全氮、全磷、有机质均高于冠层下,沙蒿群落冠层间的相应养分亦高于冠层下,而藻结皮则为冠层间低于冠层下。沿土层剖面方向,除草地表现出5—10 cm层土壤养分显著高于上层,其余群落生物结皮下伏土壤养分均随土层加深逐渐降低。苔藓结皮及藻结皮的养分富集作用主要集中在表层0—10 cm层土壤,且苔藓结皮的养分富集作用优于藻结皮。据此认为,开展旱区荒漠的生态建设时,需充分考虑植被类型同生物结皮之间的互作效应,尽量实现生物结皮、植被类型之间的最优土壤养分耦合效应。
猜你喜欢
现代园艺(2022年16期)2022-09-06
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04
现代农村科技(2021年5期)2021-12-07
土壤(2021年5期)2021-11-17
乡村科技(2021年16期)2021-10-21
中国煤炭工业(2020年1期)2020-12-28
水土保持通报(2020年6期)2020-03-10
当代陕西(2019年7期)2019-04-25
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10
散文诗世界(2016年11期)2016-11-16
