
Neuroprotection mediated by the Wnt/Frizzled signaling pathway in early brain injury induced by subarachnoid hemorrhage
2019-03-15 05:50YangWangDeJunBaoBinXuChuanDongChengYongFeiDongXiangpinWeiChaoShiNiu
中国神经再生研究(英文版) 2019年6期
Yang Wang , De-Jun Bao , Bin Xu , Chuan-Dong Cheng Yong-Fei Dong Xiang-pin Wei Chao-Shi Niu
1 Department of Neurosurgery, First Affiliated Hospital of USTC, Division of Life Sciences and Medicine, University of Science and Technology of China, Hefei, Anhui Province, China
2 Anhui Medical University Auhui Province Medical Genetic Center, Hefei, Anhui Province, China
3 Anhui Province Key Laboratory of Brain Function and Brain Disease, Hefei, Anhui Province, China
AbstractThe Wnt/Frizzled signaling pathway participates in many in flammation-linked diseases. However, the in flammatory response mediated by the Wnt/Frizzled signaling pathway in experimental subarachnoid hemorrhage has not been thoroughly investigated. Consequently, in this study, we examined the potential role of the Wnt/Frizzled signaling pathway in early brain injury in rat models of subarachnoid hemorrhage.Simultaneously, possible neuroprotective mechanisms were also investigated. Experimental subarachnoid hemorrhage rat models were induced by injecting autologous blood into the prechiasmatic cistern. Experiment 1 was designed to examine expression of the Wnt/Frizzled signaling pathway in early brain injury induced by subarachnoid hemorrhage. In total, 42 adult rats were divided into sham (injection of equivalent volume of saline), 6-, 12-, 24-, 48-, 72-hour, and 1-week subarachnoid hemorrhage groups. Experiment 2 was designed to examine neuroprotective mechanisms of the Wnt/Frizzled signaling pathway in early brain injury induced by subarachnoid hemorrhage. Rats were treated with recombinant human Wnt1 (rhwnt1), small interfering Wnt1 (siwnt1) RNA, and monoclonal antibody of Frizzled1 (anti-Frizzled1) at 48 hours after subarachnoid hemorrhage. Expression levels of Wnt1, Frizzled1, β-catenin, peroxisome proliferator-activated receptor-γ, CD36, and active nuclear factor-κB were examined by western blot assay and immuno fluorescence staining. Microglia type conversion and in flammatory cytokine levels in brain tissue were examined by immuno fluorescence staining and enzyme-linked immunosorbent assay. Our results show that compared with the sham group, expression levels of Wnt1, Frizzled1, and β-catenin were low and reduced to a minimum at 48 hours, gradually returning to baseline at 1 week after subarachnoid hemorrhage. rhwnt1 treatment markedly increased Wnt1 expression and alleviated subarachnoid hemorrhage-induced early brain injury (within 72 hours), including cortical cell apoptosis, brain edema, and neurobehavioral deficits, accompanied by increasing protein levels of β-catenin, CD36, and peroxisome proliferator-activated receptor-γ and decreasing protein levels of nuclear factor-κB. Of note, rhwnt1 promoted M2-type microglia conversion and inhibited release of in flammatory cytokines (interleukin-1β, interleukin-6, and tumor necrosis factor-α). In contrast, siwnt1 RNA and anti-Frizzled1 treatment both resulted in an opposite effect. In conclusion, the Wnt/Frizzled1 signaling pathway may participate in subarachnoid hemorrhage-induced early brain injury via inhibiting the in flammatory response, including regulating microglia type conversion and decreasing in flammatory cytokine release. The study was approved by the Animal Ethics Committee of Anhui Medical University and First Affiliated Hospital of USTC,Division of Life Sciences and Medicine, University of Science and Technology of China (approval No. LLSC-20180202) in May 2017.
Key Words: nerve regeneration; subarachnoid hemorrhage; Wnt/Frizzled signaling pathway; early brain injury; nuclear factor-κB; M2 type microglia; peroxisome proliferator-activated receptor-γ; in flammatory cytokines; neural regeneration
Introduction
Subarachnoid hemorrhage (SAH) is a familiar but severe cerebrovascular disease, resulting in neurological disorder and high mortality and disability rates (Miller et al., 2014; Okada and Suzuki, 2017). Recently, there have been significant improvements in minimally invasive and intervention therapy,but the outcome of patients who suffer a SAH still appears poor (Li et al., 2018). SAH-induced early brain injury is recognized as an early and acute cerebral vascular event that occurs within the first 72 hours, and is already regarded as a signi ficant cause of poor prognosis (Yuksel et al., 2012; Nishikawa and Suzuki, 2017). Currently, multiple pathological processes have been shown to aggravate early brain injury, including increased intracranial pressure, blood-brain barrier destruction,decrease of cerebral blood flow, in flammatory injury, and neuronal apoptosis (Yan et al., 2017; Conzen et al., 2018; Han et al., 2018). Further, the acute in flammatory response after SAH has gained increasing attention in recent years. Undoubtedly,a severe in flammatory reaction aggravates neural cell apoptosis and brain edema, and destroys the blood-brain barrier.
Immunoinflammatory responses play a critical role in early brain injury and cerebral vasospasm after SAH. After aneurysm rupture, erythrocytes, leukocytes, mononuclear macrophages, and plasma proteins are released into the subarachnoid space and in filtrate around the parenchyma. Subsequently, these in filtrated in flammatory cells activate microglia and release a large number of in flammatory cytokines (Huang et al., 2017; Shi et al., 2017; Zheng and Wong, 2017; Savarraj et al., 2018). Previous studies have shown that in flammatory factors, such as interleukin (IL)-6, IL-1β, and tumor necrosis factor alpha (TNF-α), undergo an obvious increase in the acute stage of SAH. Accordingly, interventions of IL-1 and TNF-α and their receptors can effectively improve early brain injury(Ye et al., 2016; Fan et al., 2017). Multiple signaling pathways,including Ras-MAPK-nuclear factor (NF)-κB, JAK/STAT, and TLR4/NF-κB, also participate in the release of in flammatory cytokines (Wang et al., 2014; Feng et al., 2016; Li et al., 2018).Therefore, using drug interventions to improve outcome at key points of these inflammation-associated signaling pathways has become a hot topic. Specifically, in flammation and relevant signaling pathways are important factors that lead to aggravated early brain injury after SAH. It is well known that M1-type microglia play a major role in promoting the in flammatory reaction, and can promote release of inflammatory mediators, such as TNF-α, IL-1β, and IL-6, and induce cell apoptosis, neurotoxicity, and secondary injury (Ueba et al.,2018; Zhang et al., 2018). In contrast, M2-type microglia can exert opposite and favorable effects, promoting release of anti-in flammatory mediators, and ultimately, repair of damaged arteries and tissue (Yang et al., 2018). CD36 is a member of the class B scavenger receptor family that is expressed in microglia and participates in phagocytosis. Moreover, CD36 is regulated by numerous drugs and agonists (Fang et al., 2014; Kim et al.,2017). Consequently, microglia-type and CD36 expression is recognized as a strong connection with in flammation in many diseases (Hashemi-Monfared et al., 2018). The Wnt/Frizzled signaling pathway was named because Wnt proteins are the starting signal. It is considered the most classical pathway and therefore intensely investigated. At present, research related to the Wnt/Frizzled signaling pathway has mainly focused on tumor proliferation. Hence, molecular inhibitors for specific targets of signal transduction pathways have been tested and already become anti-neoplastic drugs (Amado et al., 2014; Silva-García et al., 2014). In addition, the Wnt/Frizzled pathway is involved in a variety of related in flammatory diseases that regulate airway inflammation induced by smoke via peroxisome proliferator-activated receptor (PPAR)-δ/p38 pathways.Simultaneously, the Wnt/Frizzled pathway is suppressed, accompanied by down-regulation of PPARγ levels and up-regulation of inflammatory factors in lung tissue or bronchial epithelial cells of patients with chronic obstructive pulmonary disease caused by smoke inhalation (Kwak et al., 2015). In acute pneumonia, activation of the Wnt/Frizzled signaling pathway can reduce in flammatory damage by reducing neutrophil number and macrophage adhesion (Guo et al., 2015;Zu et al., 2016). In recent research, the Wnt/Frizzled signaling pathway was also shown to regulate proliferation and differentiation of immune cells, while a Wnt antagonist increased in flammation by promoting T cell conversion into Th2 cells.Simultaneously, Wnt3a could suppress the in flammatory response induced by pseudomonas aeruginosa, and thereby enhance macrophage phagocytosis of bacteria (Chae et al., 2016;Chen et al., 2016). These findings show that the Wnt/Frizzled pathway may modulate macrophage/microglia activation.
Lately, involvement of the Wnt/Frizzled signaling pathway in central nervous system diseases has gained increasing attention. In a cerebral ischemic model, Wnt/Frizzled pathway activation reduced infarction hemorrhage (Wang et al., 2016)and improved the microenvironment after ischemia coordinated by hypoxia-inducible factor-1α channels and vascular endothelial growth factor (Wu et al., 2016). In the acute phase of intracerebral hemorrhage, the Wnt/Frizzled pathway altered proliferation and apoptosis of neural cells and improved secondary brain injury (Zhou et al., 2014). Nevertheless,function of the Wnt/Frizzled signaling pathway in early brain injury after SAH has not yet been investigated, especially with regard to classification of microglia activation, in flammatory cytokine release, and potential mechanisms. Thus, we aimed to investigate the role of the Wnt/Frizzled signaling pathway and its underlying neuroprotective mechanisms in early brain injury after SAH in rat models, and here show involvement of the in flammatory response in this process.
Materials and Methods
Ethics and animals
A total of 150 adult male Sprague-Dawley rats aged 8 weeks were provided by the Experimental Animal Center of Anhui Medical University, China (production license No. SCXK[Wan] 2017-001; user license No. SYXK [Wan] 2017-006).
Overall, 114 rats were used to successfully establish SAH models. The average weight of healthy rats was 300—350 g.The body temperature of experimental rats was sustained at 37°C, and rats were separately fed in a constant temperature and humidity environment. All animal experiments were approved and supervised by the Animal Ethics Committee of Anhui Medical University and the First Affiliated Hospital of University of Science and Technology of China (approval No.LLSC-20180202) in May 2017. All staff abided strictly by the regulations of the National Institutes of Health concerning feeding and use of experimental animals.
SAH model induction
The prechiasmatic cistern injection method (Wang et al.,2015) was chosen to induce the SAH model. First, intraperitoneal anesthesia was accomplished using urethane (1000 mg/kg). Autologous and arterial blood was extracted from femoral arteries that were separated from femoral veins using an operating microscope. Second, a specific stereotaxic instrument was used to fix the rat's head, and a stereotaxic needle with a rounded tip and side aperture orientated towards the prechiasmatic cistern was prepared beforehand (Wang et al.,2015). The insertion position was 7.5 mm anterior to bregma in the midline, with an insertion direction along an angle of 45° in the coronal plane. The side aperture of the needle was facing the right side when the bone hole was completed. The puncture depth was approximately 10—12 mm from the skull bone surface, and the needle tip reached the optic chiasm cistern. Finally, 0.3 mL autologous and arterial blood was slowly injected into the prechiasmatic cistern over 20 seconds using an aseptic syringe pump. Correspondingly, 0.3 mL saline was injected into the prechiasmatic cistern in sham groups. Cardiac compression was performed immediately once apnea occurred after injection of blood. Most rats returned to normal respiration soon after. Vital signs of rats were observed for 45 minutes following injection of blood, and then the rats were returned to their cages at a constant temperature and humidity. Moderate normal saline was injected into subcutaneous tissue immediately to maintain homeostasis of the internal environment of experimental rats. Meanwhile, vital signs were monitored continuously and kept stable. SAH models were successfully established when 0.3 mL autologous blood was steadily injected into the prechiasmatic cistern, with brain tissue obtained at the appointed time. No rats died (0/18 rats) in the sham group. In contrast, the mortality rate of experimental rats was 27.3% (36/132 rats) after induction of SAH.
Experimental design and intervention
Experiment 1 was designed to confirm involvement of the Wnt/Frizzled signaling pathway in early brain injury after SAH. Accordingly, Wnt1, Frizzled1, and β-catenin levels were examined by western blot assay and immuno fluorescence assay at different stages. In total, 42 adult male Sprague-Dawley rats were randomly divided into seven groups (n = 6): sham,6-hour SAH, 12-hour SAH, 24-hour SAH, 48-hour SAH, 72-hour SAH, and 1-week SAH group. Subtemporal brain tissue was always covered by a blood clot in this experiment. Hence,brain tissue below the blood clot was obtained. Schematic diagram of the analyzed areas is shown in Figure 1A and B. Rats were killed by intraperitoneal anesthesia with urethane (1000 mg/kg). Brain tissue was obtained from SAH rats at different time points for western blot assay and immuno fluorescence assay. The whole experimental flow graph is shown in Figure 1C.
Experiment 2 was designed to investigate involvement of the inflammatory response including microglia somatotypes and in flammatory cytokines aggravated by early brain injury. Additionally, neuronal cells apoptosis, brain water content, and neurological behavior deficits after SAH were examined. Interventions used recombinant human Wnt1 (rhwnt1), small interfering Wnt1 RNA (siwnt1RNA), and monoclonal antibody of Frizzled1 (anti-Frizzled1). In total, 72 adult male rats were randomly divided into six groups (n = 12), including sham,SAH + control (saline), SAH + rhwnt1, SAH + siwnt1, SAH +anti-Frizzled1 + control, and SAH + anti-Frizzled1 + rhwnt1.Rats received intracerebroventricular injection of rhwnt1 and siwnt1RNA at 6 hours before they were killed. At 48 hours,neurological behavior was examined in all experimental rats.Brain tissue from the temporal lobe of six rats was cut into slices and used for terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) staining and immuno fluorescence. The remaining six rats were exsanguinated and brain samples collected for western blot assay, inflammatory cytokine level quantification, and brain edema evaluation. The whole experimental flow graph is shown in Figure 1D.
Drug treatment
Following establishment of SAH models, rhwnt1 (ab84080; Abcam Ltd., New Territories, Hong Kong, China) was dissolved and diluted in normal saline to obtain an final concentration of 1 ng/20 μL (Silva et al., 2017). Next, rhwnt1 (1 ng) was injected into the lateral cerebral ventricle using a Hamilton microsyringe guided by a stereotaxic instrument. The same volume of saline was injected into the lateral cerebral ventricle in the SAH + control group. Siwnt1RNA and control siRNA (Guangzhou RiboBio Co., Ltd., Guangzhou, China) were dissolved in RNase-free H2O to a final concentration of 1000 pmol/10 μL.Subsequently, 1000 pmol siwnt1RNA or control siRNA was diluted with transfection reagent (Entransterin vivo; Engreen,Beijing, China) and mixed evenly by shaking gently (Han et al., 2015). Following anesthesia, the mixture was intracerebroventricularly injected using a Hamilton microsyringe guided by a stereotaxic instrument. The stereotaxic coordinates were:1.5 mm posterior, 1.0 mm lateral, and 3.2 mm below the horizontal plane of bregma (Dang et al., 2015). Anti-Frizzled1 (sc-398082; Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA),a monoclonal antibody of Frizzled1, served as a Frizzled1 suppressant. Once SAH models were established, the rats received anti-Frizzled1 (3 mg/kg) by intravenous tail injection.
Western blot assay

Figure 1 Schematic of areas taken for analysis and experimental design.
In experiment 1, brain tissue samples were obtained at 6, 12,24, 48, and 72 hours and 1 week after SAH. In experiment 2, rhwnt1, siwnt1 RNA, and anti-Frizzled1 interventions were applied once SAH models were established. Brain tissue samples were all obtained at 48 hours in the different groups.Brain tissue samples were cut into small pieces using a brain chisel, and ground in a mixture of lysis buffer and phenylmethylsulphonyl fluoride (P0013; Beyotime, Shanghai, China).Protein concentrations of brain samples were measured using the Enhanced BCA Protein Assay Kit (P0009; Beyotime).First, equivalent volumes of protein samples (20 μg/lane) were heated for 5 minutes at 95°C, loaded on 12% sodium dodecyl sulphate-polyacrylamide gels, separated by electrophoresis apparatus, and electrophoretically transferred to polyvinylidene difluoride membrane (IPVH00010; Millipore, Billerica, MA,USA). Membranes were blocked with 5% bovine serum albumin for 1 hour at room temperature. Meanwhile, primary antibodies against Wnt1 (rabbit polyclonal antibody, ab85060;Abcam), Frizzled1 (goat polyclonal antibody sc-30428; Santa Cruz Biotechnology Inc.), β-catenin (rabbit monoclonal antibody, ab32572; Abcam), CD36 (rabbit polyclonal antibody, ab64014; Abcam), PPAR-γ (rabbit polyclonal antibody,ab209350; Abcam), and NF-κBp65 (rabbit polyclonal antibody, ab16502; Abcam) were diluted (1:1000), added, mixed,and incubated with membranes overnight at 4°C. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (ab9485; Abcam)was used as a loading control. Membranes were washed three times and further incubated with polyclonal donkey anti-goat IgG-horseradish peroxidase (rabbit polyclonal antibody, sc-2020; 1:5000; Santa Cruz Biotechnology Inc.) or goat anti-rabbit IgG-horseradish peroxidase (sc-2004; 1:5000; Santa Cruz Biotechnology Inc.), and conjugated secondary antibodies for 2 hours at room temperature. Finally, band signals were developed after adding enhanced chemiluminescence western reagent (P0018AM; Beyotime) and exposure to X-ray film(FF057; Beyotime). Films were scanned into images using an Epson Perfection 2480 scanner (Seiko Corp., Nagano, Japan).Related proteins were quantified with ImageJ program 1.8.0 version (NIH, Bethesda, MD, USA).
Immuno fluorescence assay
Brain tissue samples were obtained at corresponding time points after SAH. Interventions of rhwnt1, siwnt1 RNA, and anti-Frizzled1 were applied once SAH models were established. Expression of Wnt1, Frizzled1, β-catenin, and CD36 in neurons was measured by immunofluorescent double staining. CD206-positive cells were identified as M2-type microglia. First, brain samples were fixed in 4% paraformaldehyde, embedded in paraffin, and cut into 4 μm sections.Primary antibodies (1:100) of Wnt1 (rabbit polyclonal antibody, ab85060; Abcam), Frizzled1 (goat polyclonal antibody, sc-30428; Santa Cruz Biotechnology Inc.), β-catenin(rabbit monoclonal antibody, ab32572; Abcam), CD206(rabbit polyclonal antibody, ab64693; Abcam), and CD36(rabbit polyclonal antibody, ab64014; Abcam) were added and incubated for 12 hours at 4°C. Sections were washed three times, and then the corresponding secondary antibodies (1:500 dilution) were added and incubated for 1 hour.Finally, sections were covered with anti-fading mounting medium containing 4′,6-diamino-2-phenyl indole (DAPI)(C1002; Beyotime) and observed using a fluorescence microscope (BX50/BX-FLA/DP70; Olympus, Tokyo, Japan).Fluorescence intensity attached to different groups was analyzed using ImageJ program 1.8.0 version and normalized to fluorescence intensity of the sham group.
Enzyme linked immunosorbent assay assay of in flammatory cytokines
Brain tissue samples were obtained at corresponding time points after SAH. Moreover, rhwnt1, siwnt1 RNA, and anti-Frizzled1 interventions were applied once SAH models were established.Levels of IL-1β, IL-6, and TNF-α in brain tissue were measured using specific enzyme linked immunosorbent assay kits for rats(PI303, PI328, PT516; Beyotime), in accordance with the manufacturer's instructions. Brie fly, equal quantities of brain tissue from different groups were cut and ground to generate protein samples. Standard samples were diluted to different concentrations and target samples added into corresponding wells of 100 μL. Reaction wells were sealed with transparent film, and incubated at room temperature for 120 minutes. Next, prepared biotinylated antibody was added into the reaction well, and incubated for 1 hour at room temperature. Horseradish peroxidase was added to detect streptavidin, and incubated for 20 minutes at room temperature. Finally, 3,3′,5,5′-tetramethylbenzidine solution was added and incubated for 20 minutes at room temperature. After stop solution was added, absorbance values were measured at 450 nm immediately using a spectrophotometer(FilterMax F5, Molecular Devices, San Francisco, CA, USA).Values were expressed as pg/mL.
TUNEL staining
Brain tissue samples were obtained at corresponding time points after SAH. Interventions of rhwnt1, siwnt1 RNA,and anti-Frizzled1 were applied once SAH models were established. Cortical cell apoptosis was evaluated by TUNEL staining, which was performed in accordance with the manufacturer's introductions (C1086; Beyotime). Briefly,brain tissue was fixed, embedded, sliced, heated, and then dewaxed. Following dewaxing, sections were incubated with proteinase K for 15 minutes at room temperature. Next,sections were washed three times with phosphate buffered saline and then incubated with TUNEL detection liquid at 37°C for 1 hour. Sections were again washed three times.DAPI mounting medium and anti- fluorescence quenching were added and sections covered with coverslips. Finally,sections were observed using a fluorescence microscope(BX50/BX-FLA/DP70; Olympus). To measure the degree of cortical cell apoptosis, the apoptotic index was defined as the percentage of TUNEL-positive cells in each section.
Brain water content assessment
Brain tissue samples were obtained at corresponding time points after SAH. Moreover, interventions of rhwnt1, siwnt1 RNA, and anti-Frizzled1 were applied once SAH models were established. The wet/dry method was used to assess the brain water content. Following brain tissue removal and collection,brain samples were weighed immediately (wet weight), dried in an oven at 100°C for 48 hours, and then weighed again (dry weight). The brain edema index was calculated according to the formula: (wet weight - dry weight/wet weight) × 100%.
Neurological impairment
In all rats at 48 hours after intervention, activity, appetite,and neurological defects were assessed using a published and recognized scoring system to detect neurological damage (Table 1) (Yamaguchi et al., 2004; Li et al., 2018). Higher scores indicate more severe neurological impairments.

Table 1 Behavior and activity scores
Statistical analysis
All data are shown as the mean ± SEM. SPSS 11.5 software(SPSS Inc., Chicago, IL, USA) was used for statistical analysis. Data were analyzed by one-way analysis of variance followed by Scheffé F test for post hoc analysis. Statistical significance was set to α = 0.05.
Results
Decreased expression levels of Wnt1, Frizzled1, and β-catenin after SAH
To investigate expression of Wnt1, Frizzled1, and β-catenin during early brain injury after SAH, western blot assay of brain tissue was performed (Figure 2A-D). Protein levels of Wnt1 (12 hours), Frizzled1 (24 hours), and β-catenin (24 hours) were all lower compared with the sham group, being lowest at 48 hours after SAH (P < 0.05 or P < 0.01), and then gradually increasing at 72 hours (P < 0.05 or P < 0.01).Nonetheless, at 1 week after SAH, Wnt1, anti-Frizzled1,and β-catenin protein levels were still visibly lower than the sham group. Double immunofluorescence staining using Wnt1, Frizzled1, and β-catenin antibodies with a neuronal marker (NeuN) was performed (Figure 3A-F). Consistently, the results showed a similar trend as the western blot assay: expression of Wnt1, Frizzled1, and β-catenin attached to the cytoplasm of neuronal cells reached a lowest point at 48 hours. The fluorescent intensity of Wnt1, Frizzled1, and β-catenin returned to normal levels at one week. These results suggest that the Wnt/Frizzled signaling pathway may participate in the pathological process during early brain injury, and is apparently suppressed after SAH. Further, 48 hours after SAH might be the most appropriate time point for intervention in experiment 2.
Rhwnt1 treatment increases and siwnt1 treatment decreases Wnt1 protein levels, while anti-Frizzled1 treatment decreases Frizzled1 protein levels in brain tissue under SAH conditions

Figure 2 Changes in protein levels of Wnt1, Frizzled1, and β-catenin measured by western blot assay after SAH.

Figure 3 Double immuno fluorescence of Wnt1,Frizzled1, and β-catenin in neurons after experimental SAH.
Western blot assay (Figure 4A-D) and immuno fluorescent staining (Figure 4E-H) both showed significantly lower Wnt1 protein levels at 48 hours after SAH compared with the sham group (P < 0.05). Wnt1 protein levels were signi ficantly increased by rhwnt1 intervention and significantly decreased by siwnt1 RNA intervention (P < 0.01). Further,western blot assay and immuno fluorescent staining results also indicated down-regulated Frizzled1 protein levels in the SAH + control group (P < 0.01), which were significantly further reduced by anti-Frizzled1 treatment (P < 0.01).
Rhwnt1 treatment decreases, while siwnt1RNA and anti-Frizzled treatment increases SAH-induced cortical cell apoptosis
TUNEL staining was performed to examine the effect of the rhwnt1, siwnt1, and anti-Frizzled1 interventions on cortical cell apoptosis in the brain at 48 hours after SAH (Figure 5A and B). Significant increase of TUNEL-immunoreactive cells was observed in the SAH + control group compared with the sham group (P < 0.01). The percentage of TUNEL-immunoreactive cells was decreased by rhwnt1 treatment (P <0.01) and increased by siwnt1 and anti-Frizzled1 treatment(P < 0.05). Simultaneously, the pro-apoptotic effect of anti-Frizzled1 was partially neutralized by rhwnt1 (P < 0.05).

Figure 4 Changes in Wnt1 and Frizzled1 expression altered by rhwnt1, siwnt1, and anti-Frizzled1 treatment after experimental SAH.

Figure 5 Effect of rhwnt1, siwnt1,and anti-Frizzled1 treatment on cortical cell apoptosis and brain edema after experimental SAH.
Rhwnt1 treatment decreases, while siwnt1RNA and anti-Frizzled1 treatment increases SAH-induced brain edema and neurological impairment
First, brain edema index was significantly higher in the SAH + control group than the sham group (P < 0.01) (Fig-ure 5C). In contrast, after rhwnt1 treatment, brain edema index was significantly less pronounced (P < 0.01). Mean brain edema index was higher in rats after siwnt1RNA and anti-Frizzled1 treatment than in the SAH + control group(P < 0.05). Simultaneously, brain water content was signi ficantly lower in the SAH + rhwnt1 group than in the SAH +control group (P < 0.01). However, there were no significant variations between the SAH + anti-Frizzled + control group and SAH + anti-Frizzled1 + rhwnt1 group. Rats with SAH showed severe neurological behavioral defects compared with the sham group (2.87 ± 0.32 points vs. 0.67 ± 0.15 points; P <0.01; Figure 5D). These neurological behavioral defects were significantly less pronounced after rhwnt1 treatment (2.03 ±0.58 points vs. 2.87± 0.32 points; P < 0.01). After siwnt1RNA(3.35 ± 0.29 points vs. 2.87 ± 0.32 points) and anti-Frizzled1(3.41 ± 0.47 points vs. 2.87 ± 0.32 points) treatment, neurological behavioral defects were significantly more pronounced compared with the SAH + control group (P < 0.05). Moreover, remarkable variation in neurological behavioral defects was detected in the SAH + anti-Frizzled1 + rhwnt1 group compared with the SAH + anti-Frizzled + control group (3.03± 0.38 points vs. 3.41 ± 0.47 points; P < 0.05).
Rhwnt1 promotes M2 microglia conversion and inhibits in flammatory cytokine release, while siwnt1 and anti-Frizzled1 have opposite effects after experimental SAH
The effect of rhwnt1, siwnt1, and anti-Frizzled1 interventions on microglia type in the brain at 48 hours after SAH was investigated by immunofluorescence staining with CD206(Figure 6A and B). Significant reduction of CD206-immunoreactive cells was observed in the SAH + control group compared with the sham group (P < 0.05). The percentage of CD206-immunoreactive cells was increased by rhwnt1 treatment and decreased by siwnt1RNA and anti-Frizzled1 treatment (P < 0.05 or P < 0.01). However, no remarkable difference in CD206-immunoreactive cells was detected between the SAH + anti-Frizzled1 control group and SAH +anti-Frizzled1 + rhwnt1 group. In the SAH + control group,average levels of IL-1β, IL-6, and TNF-α in brain tissue were significantly increased compared with the sham group (P <0.01; Figure 6C). Rhwnt1 treatment abolished this increase (P< 0.05), while siwnt1RNA and anti-Frizzled1 treatment intensified the increase (P < 0.05). The proin flammatory effect of anti-Frizzled1 was partially, but significantly, neutralized by rhwnt1 treatment (P < 0.05).
Rhwnt1 treatment increases β-catenin, PPAR-γ, and CD36 expression and decreases NF-κBp65 expression,while siwnt1 and anti-Frizzled1 treatment exert opposite effects after experimental SAH
Expression of NF-κBp65, β-catenin, CD36, and PPAR-γ was examined to determine the mechanism of microglia type conversion by western blot assay (Figure 7A-E) and immuno fluorescent double labeling (Figure 8A-D). Evident reduction in protein levels of β-catenin, CD36, and PPAR-γ was found in the SAH + control group (P < 0.01). Rhwnt1 treatment resulted in significantly higher protein levels of β-catenin, CD36, and PPAR-γ compared with the SAH +control group (P < 0.01). However, β-catenin, CD36, and PPAR-γ expression was less after siwnt1RNA and anti-Frizzled1 intervention compared with the SAH + control group(P < 0.05, P < 0.01). These results also showed that protein levels of β-catenin, CD36, and PPAR-γ were lower after anti-Frizzled1 treatment compared with the siwnt1RNA group(P < 0.05 or P < 0.01). In contrast, a significant increase in protein levels of NF-κBp65 was observed in the SAH + control group (P < 0.05), while rhwnt1 treatment significantly down-regulated NF-κBp65 protein levels (P < 0.01). NF-κBp65 was significantly higher in the SAH + anti-Frizzled1 control group than the SAH + control group (P < 0.05).
Discussion
Experiment 1 showed decreased levels of Wnt1, Frizzled1,and β-catenin in brain tissue and neurons after SAH, with the most noticeable inhibition time point being 48 hours after SAH. In experiment 2, we found the following results: first,rhwnt1 treatment up-regulated the SAH-induced decrease in Wnt1, β-catenin, PPAR-γ, and CD36, and further down-regulated the SAH-induced decrease in NF-κBp65 in brain tissue and neurons. Meanwhile, siwnt1RNA had a down-regulatory effect. Anti-Frizzled1 inhibited levels of Frizzled1, β-catenin,PPAR-γ, and CD36, and enhanced NF-κBp65 in brain tissue and neurons, but had no effect on Wnt1 levels. Anti-Frizzled1 effects were partially reversed by rhwnt1 treatment. Further,rhwnt1 treatment alleviated brain edema, cortical apoptosis,and impaired neurological behavior after experimental SAH,while siwnt1RNA and anti-Frizzled1 had an aggravated effect that was partially reversed by rhwnt1 treatment. Ultimately,rhwnt1 treatment promoted microglia conversion into M2-type, and inhibited in flammatory cytokine release. Again, siwnt1RNA and anti-Frizzled1 exerted an opposite effect. Based on the above results, we hypothesized that decreased Wnt1 protein secretion and expression occurs after SAH, which is accompanied by suppression of the Wnt/Frizzled signaling pathway. Extracellular Wnt1 did not interact with Frizzled1,which was mediated by β-catenin accumulation inside the cell. Decreased β-catenin content in the nucleus decreased PPAR-γ expression, which combined with the transcription factor, TCF/LEF. Alternatively, reduction of PPAR-γ expression down-regulated expression of the membrane receptor,CD36, and promoted conversion of microglia to a M2 classification. Since low concentrations of intranuclear PPAR-γ were unable to effectively antagonize NF-κB, relevant in flammatory signaling pathways were activated. Consequently,release of a large number of in flammatory factors was inevitable, which directly caused nerve cell damage and exacerbated early brain injury (hypothesis figure of in flammation-induced neural damage mediated by Wnt/Frizzled1 signaling pathway suppression after SAH; Figure 9).
It is generally known that the in flammatory reaction is an important host defense reaction to brain injury after SAH.Accumulative evidence has shown that excessive in flammatory responses following SAH aggravate brain injury, including blood-brain barrier disruption, brain edema aggravation,and neuronal cell death (Guo et al., 2016; Chaudhry et al.,2017; Frontera et al., 2017; Yuan et al., 2018). Activation of in flammation following SAH results from microglial activation, perihematomic inflammation reaction, infiltration of blood-derived inflammatory cells, and release of inflammatory cytokines (including IL-1β, IL-6, and TNF-α) (Yin et al.,2017; Schneider et al., 2018). At present, the Wnt/Frizzled signaling pathway is strongly associated with common inflammatory diseases. For example, it is highly expressed in a mouse model of chronic asthma as well as human patients,and regulates development of airway remodeling in chronic asthma. Blocking this pathway dampens airway remodeling by down-regulating TGF-β and tenascin C/PDGFR, resulting in subepithelial fibrosis and smooth muscle hyperplasia (Kwak et al., 2015). Recently, investigators have suggested that the Wnt/Frizzled signaling pathway might play important roles in different central nervous system diseases (Sorcini et al.,2017). These findings demonstrate that TWS119, an inhibitor of glycogen synthase kinase 3β (a key node in the Wnt/Frizzled signaling pathway), reduced recombinant tissue plasminogen activator-induced hemorrhagic transformation and attenuated blood-brain barrier disruption, possibly through activating the Wnt/Frizzled signaling pathway by increasing β-catenin, claudin-3, and ZO-1 protein expression and suppressing glycogen synthase kinase 3β expression (Wang et al.,2016). Another study showed that galangin might function as a multi-target drug for treating ischemic stroke by improving the microenvironment of neurovascular units. This included ameliorating neurological defects and neurovascular unit damage after middle cerebral artery occlusion via activating Wnt/β-catenin and hypoxia-inducible factor-1α/vascular endothelial growth factor signaling pathways (Wu et al., 2016).In addition, the Wnt/β-catenin signaling pathway is relevant to cell apoptosis and expression of proliferating cell nuclear antigen in rat brain, regulating the balance between cell apoptosis and cell proliferation following intracerebral hemorrhage induction (Zhou et al., 2014; Ma et al., 2018).
PPAR-γ is a ligand-dependent transcription factor that can regulate expression of the scavenger receptor, CD36,which is of significance for phagocytic activity and microglial phenotypes (Tyagi et al., 2011; Li et al., 2015; Zhao et al.,2015; Flores et al., 2016). NF-κB is a transcription factor that modulates production of many pro-inflammatory enzymes,chemokines, and adhesion molecules, resulting in massive amplification of the secondary inflammatory response after cerebral stroke (Gan et al., 2016; Peng et al., 2018; Yue et al.,2018). In both in vitro and in vivo experiments, agonists of PPAR-γ as therapeutic targets, decrease expression of pro-inflammatory determinants (including IL-1β, TNF-α, matrix metalloproteinase-9, and inducible nitric oxide synthase) via interaction with the transcription factor, NF-κB, and inhibiting DNA binding of NF-κB (Wu et al., 2016). In addition,TAK-242, a TLR4 inhibitor, remarkably relieved inflammatory injury after intracerebral hemorrhage, and improved neurological deficits by up-regulating CD36 expression in microglia, increasing phagocytic capacity of microglia, and improving hematoma absorption in intracerebral hemorrhage mice (Fang et al., 2014). At present, the internal relationship between the Wnt/Frizzled signaling pathway, PPAR-γ,CD36, NF-κB, and in flammatory cytokine release is still not elaborated in experimental SAH. Nevertheless, in this study,reduced PPAR-γ expression directly led to down-regulation of CD36 and phagocytic capacity of microglia, and was associated with suppression of the Wnt/Frizzled signaling pathway after SAH. Alternatively, activation of the Wnt/Frizzled signaling pathway generated an opposite effect.
This current study has some limitations. First, we have not determined the precise mechanism of the Wnt/Frizzled signaling pathway in regulating PPAR-γ expression. In addition, it is not possible to accurately evaluate microglia type conversion using immuno fluorescence, and flow cytometry is more suitable. Nevertheless, in conclusion, our study provides comprehensive and adequate evidence supporting the potential effects of Wnt/Frizzled on an experimental SAH rat model and its underlying neuroprotective mechanisms.First, rhwnt1 ameliorated protein expression of key nodes(specifically, Wnt1, Frizzled1, and β-catenin), as well as brain edema, cortical cell apoptosis, and neurological defects induced by SAH. Siwnt1RNA and anti-Frizzled treatment had an opposite effect. Moreover, rhwnt1 promoted microglia conversion into M2-type and inhibited inflammatory cytokine release via regulation of PPAR-γ, CD36, and NF-κBp65 expression. All data in these experiments confirm that the neuroprotective effect of the Wnt/Frizzled signaling pathway might result from inhibition of inflammatory responses after SAH. Therefore, the Wnt/Frizzled signaling pathway may function as a multi-target in the treatment of hemorrhagic stroke.
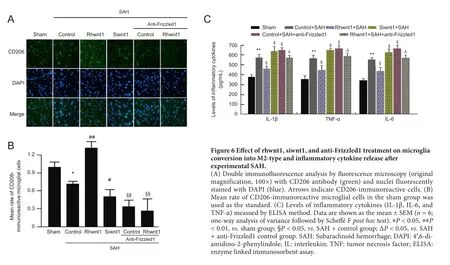
Figure 6 Effect of rhwnt1, siwnt1, and anti-Frizzled1 treatment on microglia conversion into M2-type and in flammatory cytokine release after experimental SAH.
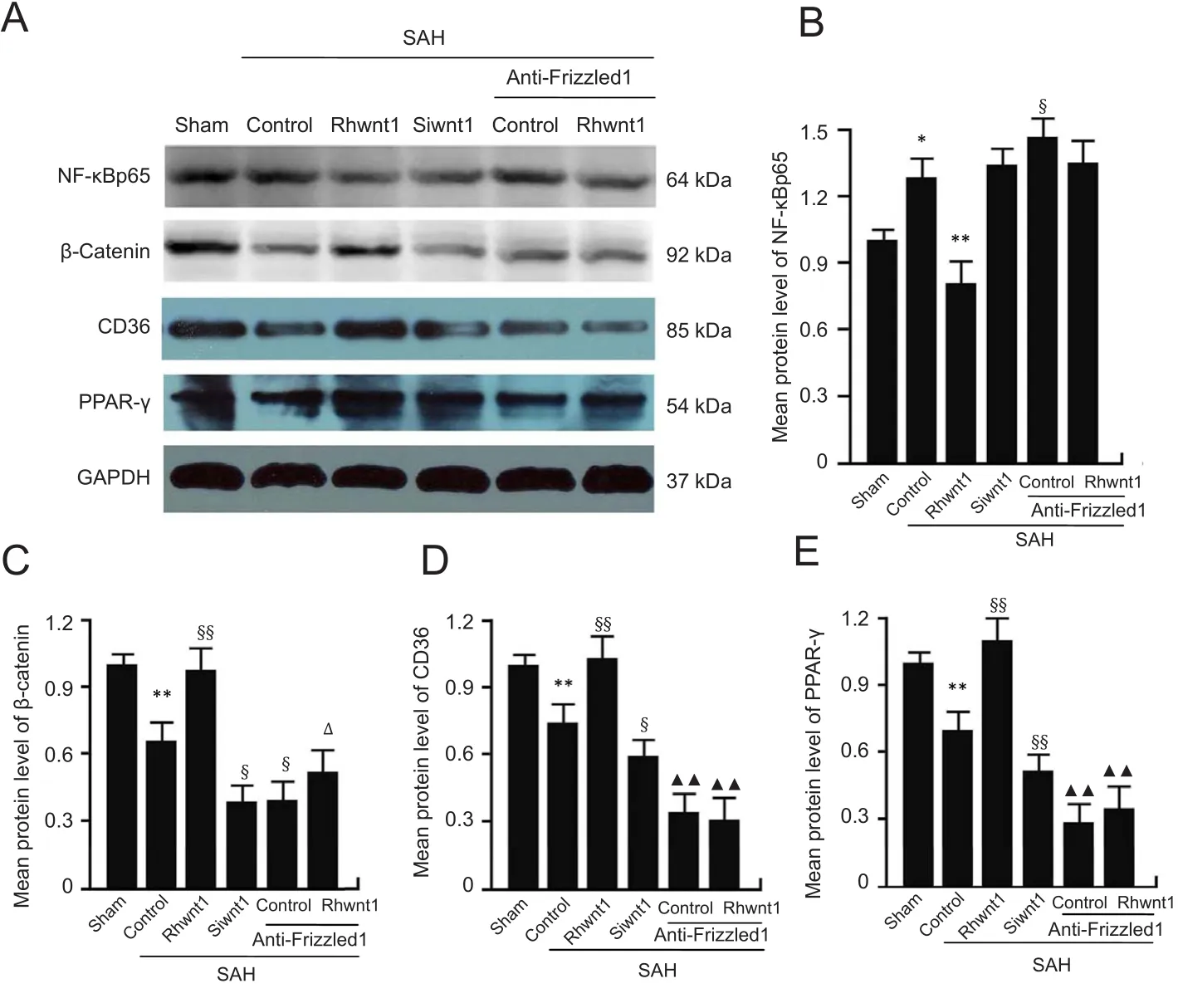
Figure 7 Changes in NF-κBp65,β-catenin, CD36, and PPAR-γ following rhwnt1, siwnt1, and anti-Frizzled1 intervention after experimental SAH.

Figure 8 Changes in β-catenin and CD36 following rhwnt1, siwnt1, and anti-Frizzled1 intervention after experimental SAH.

Figure 9 Schematic showing the role of the Wnt/Frizzled signaling pathway and potential mechanisms of the neuroprotective effect on early brain injury.
Author contributions:Study conception and design, quality assurance and control: XPW and CSN; experimental implementation and paper writing: YW and DJB; analytic strategy design: BX; literature review and preparation of Materials and Methods section: CDC; manuscript reviewing and editing: YFD.All authors read and approved the final version of the manuscript.
Conflicts of interest:The authors declare that there is no conflict of interests regarding the publication of this paper.
Financial support:This study was supported by the Natural Science Foundation of Anhui Province of China, No. 1508085QH184 (to YW). The funding body played no role in the study design, in the collection, analysis and interpretation of data, in the writing of the paper, and in the decision to submit the paper for publication.
Institutional review board statement:The study was approved by Animal Ethics Committee of Anhui Medical University and First Affiliated Hospital of USTC, Division of Life Sciences and Medicine, University of Science and Technology of China (approval No. LLSC-20180202) in May 2017.
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix, tweak,and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
- 中国神经再生研究(英文版)的其它文章
- Busting the myth: more good than harm in transgenic cells
- Comparative study of microarray and experimental data on Schwann cells in peripheral nerve degeneration and regeneration: big data analysis
- Lessons from glaucoma: rethinking the fluid-brain barriers in common neurodegenerative disorders
- Characteristics and advantages of adenoassociated virus vector-mediated gene therapy for neurodegenerative diseases
- Gene expression changes in dorsal root ganglia following peripheral nerve injury: roles in in flammation, cell death and nociception
- Nicotinamide adenine dinucleotide phosphate oxidase activation and neuronal death after ischemic stroke