
鸡公山落叶阔叶林藤本植物寄主选择与分布规律
2019-02-28 05:44:14琚煜熙王忠伟刘彦春
生态与农村环境学报 2019年2期
琚煜熙,尚 晴,王忠伟,刘彦春
(1.信阳市鸡公山国家级自然保护区管理局,河南 信阳 464133;2.黄河水利职业技术学院,河南 开封 475004;3.河南大学生命科学学院,河南 开封 475004)
与农田和草地生态系统不同,森林具有明显的水平和垂直结构[1]。其垂直结构表现为在垂直方向上可区分为明显的3个层次,即乔木层、灌木层和草本层[2]。此外,在森林垂直方向上有1类物种,即藤本植物,又称为层间植物,贯穿于森林的3个层次[3-4]。作为热带和亚热带地区森林生态系统中的附生类植物,藤本植物对于生态系统的结构与功能的维持具有重要作用[5]。研究表明,由于人为干扰及气候变化的综合影响,全球尺度上藤本植物的密度和生物量呈增长趋势[6]。藤本植物在生态系统中的作用表现在2个方面:一是藤本植物通过截获养分、水分和光照对树木生长产生直接竞争[7]。PÉREZ-SALICRUP等[8]研究显示藤本植物通过对地上和地下的资源竞争导致枫香树(Liquidambarstyraciflua)光合作用和生物量分配均显著下降。藤本植物依附林木生长的过程中通过气生根和缠绕等方式对林木树皮、树干产生强烈的机械破坏,甚至会绞杀林木,造成树木的死亡,因此会降低森林的生产力[9]。在森林火烧迹地和采伐迹地藤本植物会迅速侵入并占据林地,从而降低林木幼苗的生长、更新和存活率,这种对树木的负面影响会显著改变森林的结构、组成和动态[10]。二是藤本植物贯穿于森林内的不同层次,是很多小型动物(如松鼠和蜥蜴)和昆虫的栖息地及林内上下活动的通道;藤本植物在未找到依附林木而在地表匍匐生长时对于减少水土流失、加快凋落物分解具有重要积极作用,如LIU等[11]研究发现藤本植物在地表匍匐阶段有助于改善土壤的养分含量。
鉴于藤本植物结构和功能的特殊性,针对其在森林生态系统中作用的研究得到广泛关注[4,12-14]。然而,目前就藤本植物寄主选择和分布状况的研究甚少。笔者以我国华中鸡公山地区典型落叶阔叶林为研究对象,对其中的藤本植物进行调查(以吸盘类藤本为主),以期解决以下问题:(1)藤本植物在选择依附树种时是否具有选择性?(2)藤本植物在树干表面向上生长时,是否发生分布方向的改变?
1 材料与方法
1.1 研究区概况
研究地点位于河南省信阳市鸡公山国家级自然保护区(31°46′~31°51′ N,114°01′~114°06′ E),相对海拔高度120~810 m,主峰篱笆寨海拔811 m,属中山类型。该地区的年均温度为15.2 ℃,1月份均温为0.6 ℃,7月份均温为24.7 ℃,年均降水量1 098 mm,大多数集中分布于4—8月,年均蒸发量1 378.8 mm。土壤类型以黄棕壤为主。该地区是我国北亚热带—暖温带过渡区域,地带性植被类型为落叶阔叶林和常绿落叶阔叶混交林。但是原生植被早已不存在,现在大多数为次生林。主要的次生林类型有麻栎(Quercusacutissima)-栓皮栎(Quercusvariabilis)-枫香(Liquidambarformosana)混交林,麻栎-马尾松(Pinusmassoniana)混交林。优势灌木物种主要有黄荆(Vitexnegundo),山胡椒(Linderaglauce)和三角枫(Acerbuergerianum)等。林下藤本
植物丰富,是构成和维持该地区森林生态系统物种多样性的重要成分。主要的藤本物种有络石(Trachelospermumjasminoides),常春藤 (Hederamepalensisvar.sinensis),南蛇藤(Celastrusorbiculatus)和绞股蓝(Gymostemmapentaphyllum)等。
1.2 样木选设
在鸡公山国家级自然保护区内选择坡向(东偏南15°)和坡度(20°~25°)变幅较小的集水区(海拔210 m),区内的森林类型是该地区典型植被—落叶阔叶林,优势乔木树种是麻栎和枫香,其中零星分布少量的马尾松;灌木树种主要是山胡椒,三角枫和朴树等;面积约3 hm2。该林分密度为1 033.9株·hm-2,林分平均胸径为(21.4±4.8) cm,林分平均林龄为52 a,林分盖度为87%。
在该林分内先按随机数法确定样点,再以样点为中心确定目标样木。以该林分的坡下位置为起点,向林内每隔一定距离确定一个样点,以该样点为中心,直径3 m范围内选择有藤本植物附生的麻栎、枫香和马尾松为调查样木。由于同时具备符合条件的3个乔木树种的样点不多,多数样点周围只有2棵着生有藤本植物的乔木,为获得足够样本量的调查样木(各25棵),按随机原则从山脚下向上沿近似直线方向共设置32个样点(相邻样点距离10~15 m, 随机数产生)。将每株树作为研究个体,调查附生于树干上的藤本植物数量。
1.3 测定指标与方法
(1)样木调查:记录每株树的物种名、表面有无苔藓着生、树皮是否粗糙等基本特征,利用胸径尺测定其胸径(距地面130 cm)和基径(距地面5 cm)。
(2)藤本植物调查:将树干表面沿平行于山体等高线的方向区分为坡上位置与坡下位置2个方位,调查基径处(5 cm)和胸径(130 cm)处树干的2个方位上着生的藤本植物状况。记录藤本植物名、密度(在每个方位上着生的藤本数量)和生长状况(死亡或存活)。
1.4 数据处理
将所有野外调查数据录入Microsoft Excel 2010软件,按树种、有无苔藓覆盖和坡向方位等进行汇总计算,求出平均值与标准误差。采用One-way ANOVA和多重比较(LSD)检验树种、苔藓和方位对藤本植物分布影响的显著性。采用一元线性回归分析不同树种藤本植物着生数量与胸径大小的关系。统计分析在SPSS 16.0软件中完成。
2 结果与分析
2.1 树种对藤本植物分布的影响
如图1所示,在树干基径部位,枫香和麻栎表面附生死亡的藤本植物数量1.59株,显著高于马尾松树表面附生的死亡藤本植物(P<0.05)。此外,枫香和麻栎表面附生的活藤本植物数量均为4.19株,也显著高于马尾松表面附生的活藤本植物(P<0.05)。由于死亡及活藤本数量在不同树种之间的显著差异,导致枫香和麻栎表面总的藤本植物数量(5.99和5.79株)显著高于马尾松(P<0.05)。然而,无论是死亡还是活藤本植物在枫香和麻栎之间均无显著差异。
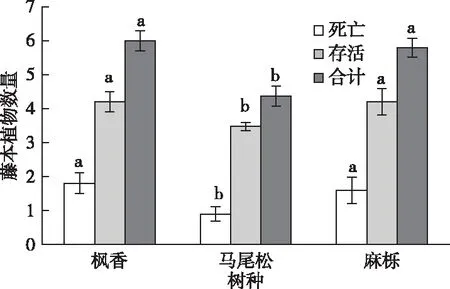
同一颜色直方柱上方英文小写字母不同表示不同树种间藤本植物数量差异显著(P<0.05)。
2.2 树干表面苔藓状况对藤本植物的影响
枫香、麻栎和马尾松树干表面附生苔藓的分别占总样木的64%、56%和71%。无论是区分树种还是综合分析,苔藓有无均对藤本植物分布无显著影响。如图2所示,有苔藓着生的树干表面的藤本数量为5.54株,高于无苔藓着生的树干(5.18株),但是未达显著差异(P>0.05)。有苔藓覆盖的树干表面活的藤本植物数量为4.15株,略高于无苔藓覆盖的树干;而死亡藤本植物的数量则略低于后者,但是均未达显著水平(P>0.05)。
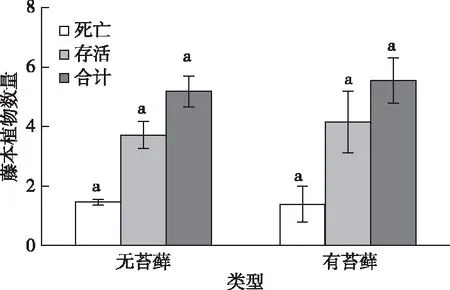
同一颜色直方柱上方英文小写字母不同表示有无苔藓间藤本植物数量差异显著(P<0.05)。
2.3 坡位对树干不同高度处藤本植物分布的影响
不同坡位藤本植物在树干表面的分布见图3。
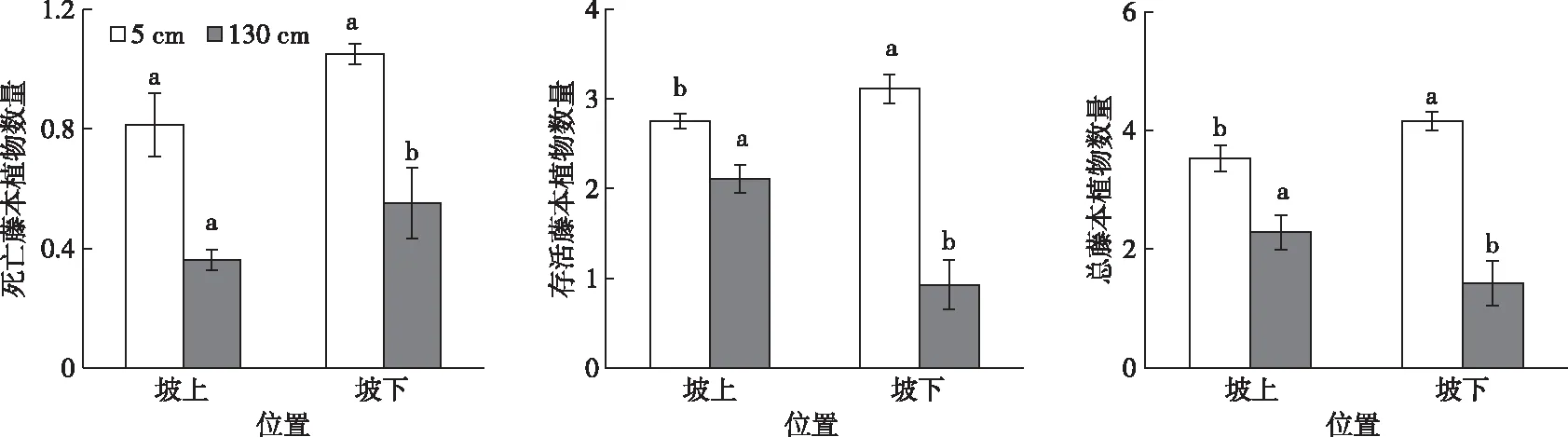
同一颜色直方柱上方英文小写字母不同表示不同位置间藤本植物数量差异显著(P<0.05)。n=75。
树干坡下位表面的死亡藤本植物在胸径位置为0.55株,显著高于树干坡上位表面的数量(P<0.05);然而,基径位置(5 cm)处的死亡藤本植物,在坡上位与坡下之间无显著差异(P>0.05)。活藤本植物在树干基径处坡下位的数量(3.12株)显著高于坡上位(P<0.05);与基径处相反,在胸径位置处树干坡上位的藤本数量(2.11株)显著高于树干坡下位(P<0.05)。就藤本总数量而言,树干坡上位的藤本植物数量(3.53株)显著低于坡下位(P<0.05)。然而,在树干胸径位置(130 cm)坡上位置的藤本植物数量为2.28株,显著高于坡下位(P<0.05)。
2.4 胸径大小对藤本植物密度的影响
由图4可知,枫香与麻栎树干着生的藤本植物数量与其胸径大小呈显著正相关(R2=0.283 6,P<0.01;R2=0.210 4,P<0.01)。对于针叶树种马尾松而言,其树干着生的藤本植物数量与胸径呈负相关关系(R2=0.170 7,P<0.05)。
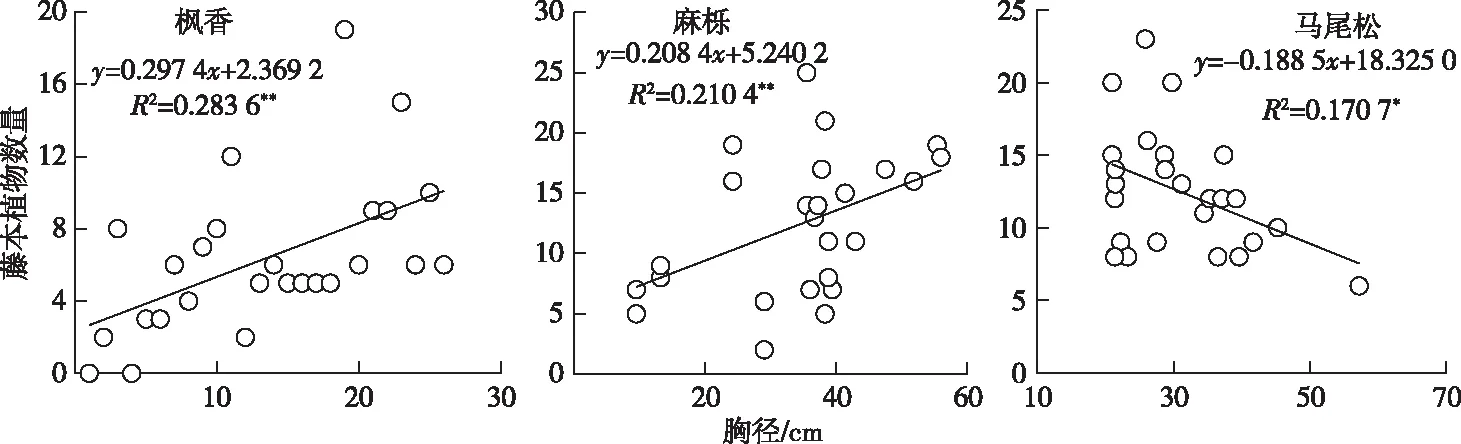
** P<0.01; *P<0.05。
3 讨论
3.1 藤本植物对树种的选择性
由于缺乏发达的自我支撑系统,为了获取更多的光资源,藤本植物必须依附于其他高大的乔木或灌木以爬到林木的顶端,并迅速扩展和占领林冠层[15-16]。在攀爬过程中,藤本植物会通过缠绕、吸附、卷须和倒刺等方式攀缘于可依附的林木上。绝大多数藤本植物植根于土壤中,其生长发育要经历地面匍匐到依附林木直立生长的过程,通常认为藤本植物寻获依附林木的机理是光调节机制。然而,其在选择依附林木方面是否具有选择性则鲜有报道[9]。笔者在鸡公山开展调查时发现藤本植物在阔叶树种(枫香和麻栎)上的附生数量显著高于针叶树种(马尾松),在一定程度上证实了藤本植物对树种的选择性。导致这一现象的原因可能有:首先,阔叶树种在养分的周转循环方面比针叶树更具优势,其树皮老化分解释放的养分更易于藤本植物的吸收;针叶树富含脂类等难分解成分,其释放的养分不易于被藤本植物根系吸收;其次,从树干表面形态特征来看,针叶树(如马尾松)表面的树皮会不定期地片状脱落,不利于藤本植物的吸附。再次,该研究选择的林分以阔叶树为主,尽管采用随机原则进行调查,着生藤本植物较多的阔叶树比针叶树出现的机率更高。
3.2 树干微环境对藤本分布的影响
研究还发现藤本植物在依附于寄主乔木生长时会发生依附部位的转变,即在树干的地表基径处以坡下位为主;而在胸径处以坡上位为主(图3),这种分布的转变显示了藤本植物生长过程中对环境条件的响应与适应。研究调查的林分具有一定的坡度,藤本植物本身无支撑结构,为争取更多的光照资源,其高生长会借助地势从坡下向坡上的方向生长[17],在匍匐生长过程中遇到林木而依附,因此,树干接近地表位置处坡下位会着生更高密度的藤本植物。当藤本植物捕获寄主林木之后,藤本植物会继续沿树干攀爬向上生长。在山地森林中,白天风向通常是从山谷吹向山脊,一方面,藤本植物顶芽在风力的作用下逐渐发生生长方向的转变,沿树干表面长向背风的一面;另一方面,风会提高水分的散失,树干迎风面方向转变较慢的藤本植物可能更易于在风害中死亡。该研究中的3个乔木树种表面大多数均生长有苔藓,苔藓的存在可改善树干表面的水分和养分状况。与研究预期相反,笔者并未发现藤本植物对有苔藓覆盖的树木具有更高的偏好。
3.3 树干大小对藤本密度的影响
研究发现,藤本植物密度与寄主树干胸径大小呈显著相关关系,阔叶树种枫香和麻栎为正相关。一项在亚马逊流域Bolivia半落叶季雨林中开展的研究发现,相比小径级树木,大径级树木能着生更多的藤本植物[7]。然而,这种关系受到寄主树木周边环境的影响。例如,当林分中胸径介于10~30 cm的林木个体比例很高时这种正相关关系就会减弱。大径级阔叶树附生更高的藤本植物可归因于2点:一方面,大径级林木的树龄较大,其被藤本寄生的机率提高了;另一方面,大径级林木具有更高的表面积,有利于藤本植物着生。与阔叶树不同的是,针叶树种马尾松表面着生的藤本植物密度与胸径大小呈负相关。这可能与马尾松树皮脱落特性有关,大径级马尾松树皮皲裂更显著,脱落更明显,不利于藤本植物着生。因此,森林中树种及胸径大小的差异均是影响森林中物种多样性[18-20],尤其是附生植物生存与维持的重要因子。
4 结论
通过对鸡公山地区落叶阔叶林中藤本植物的寄主选择性和分布规律的调查发现,藤本植物在依附林木进行直立生长的过程中对树种选择表现出一定程度的偏好,即更多选择阔叶树种,并且大径级阔叶树具有更高藤本植物密度。这种选择性与乔木本身树皮状况、养分可利用性和周边微环境有关。此外,研究还发现藤本植物在树干上的分布会从树干迎风面逐渐转向到背风面,以获得最大的生存和生长机会。该研究结果有助于提高对藤本植物在森林生态系统中生存和生长策略的理解和认识,为维持森林生态系统物种共存及多样性提供科学指导。
猜你喜欢
疯狂英语·新读写(2023年10期)2023-11-22 03:31:30
广东蚕业(2022年4期)2022-11-23 13:31:55
乡村地理(2022年2期)2022-05-30 10:48:04
军事文摘(2021年18期)2021-12-02 01:28:36
环境与生活(2021年5期)2021-06-24 18:09:52
湖北农机化(2020年16期)2020-10-21 01:29:08
江西农业(2020年13期)2020-09-08 07:44:54
儿童故事画报·自然探秘(2017年7期)2018-03-14 19:05:32
当代贵州(2016年23期)2016-07-21 08:55:34
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18 10:59:56
