
太湖沉积物反硝化功能基因丰度及其与N2O通量的关系
2019-02-26 02:46刘德鸿文帅龙龚琬晴钟继承钟文辉
生态环境学报 2019年1期
刘德鸿 ,文帅龙,龚琬晴,钟继承*,钟文辉
1. 南京师范大学地理科学学院,江苏 南京 210023;2. 湖泊与环境国家重点实验室/中国科学院南京地理与湖泊研究所,江苏 南京 210008;3. 洛阳市共生微生物与绿色发展重点实验室/河南科技大学农学院,河南 洛阳 471023
随着人类社会的快速发展,大量氮素进入生态系统,打破了原有生态系统氮循环的平衡,导致一系列环境问题的发生。湖泊生态系统氮循环作为生态系统的重要组成部分,其重要性不容忽视,尤其是近年来,湖泊富营养化程度的加剧,已成为氧化亚氮(N2O)排放研究的新热点(Yao et al.,2016)。而反硝化作用作为生态系统氮循环中重要的组成部分,早在上个世纪 70年代已受到研究者的广泛关注(Koike et al.,1978)。
反硝化过程产生的N2O逐年增多(Bouwman et al.,2013),加剧温室效应,有关反硝化的机制、影响因素以及N2O排放及机制等研究,在农田(李飞跃等,2012)、森林(Groffman et al.,1989)、海洋(Singh et al.,2010)、湿地(Burgin et al.,2015)、湖泊(Myrstener et al.,2016)等生态系统中被广泛开展。反硝化作用主要由反硝化微生物驱动完成,依次经过硝酸还原酶、亚硝酸还原酶,一氧化氮还原酶以及氧化亚氮还原酶催化,最终被还原为N2,对应基因编码分别为:narG或napA,nirS或nirK,norB及nosZ。反硝化微生物群落结构及功能与外界环境因子密切相关,研究发现,氮素含量和pH对反硝化细菌丰度有显著影响(鲍林林等,2016),沉积物有机质与nirS丰度呈正相关(Hou et al.,2013),反硝化细菌丰度存在明显的季节变化(Lisa et al.,2014),添加生物炭的土壤N2O排放与nosZ关系密切(Cayuela et al.,2013),N2O排放量随施用硝态氮量增加而显著增加,但与反硝化基因丰度之间并没有显著相关性(郑燕等,2012)。因此,反硝化功能微生物群落结构和丰度及其对环境因子的响应与N2O排放的关系,由于生态环境的不同而存在较大差异。为此,本研究选取太湖不同生态类型湖区沉积物为研究对象,分析了不同季节沉积物化学性质、N2O通量以及反硝化功能基因丰度,并探讨了它们之间的关系,旨在探明太湖沉积物反硝化功能基因丰度的季节变化,及其与环境因子的关系,初步揭示影响太湖沉积物N2O排放的微生物学机理,为进一步深入研究太湖沉积物N2O排放机制提供数据支持和理论基础。
图1 太湖采样点位分布图Fig.1 Distribution of sampling sites in Taihu Lake
1 材料方法
1.1 采样区域概况
太湖是中国第三大淡水湖,地处亚热带,气候温和湿润,属季风气候,湖水面积2338 km2,平均水深 1.9 m,是典型的大型浅水型湖泊,其位于经济发达的长三角地区,水体具有供水、渔业、航运、旅游等重要功能,对该区域经济发展具有重要作用,受人为活动影响很大,过量氮磷的输入导致水体持续富营养化(Xu et al.,2016)。太湖有着复杂的生态类型,具有草型、藻型及敞水区。采样点S1位于梅梁弯为典型的藻型湖区,一年中多数月份蓝藻占绝对优势,平均水深约3 m,无沉水植物,底部沉积有较多的淤泥,是太湖富营养化最严重的湖区;S2位于中部较开阔区域,无大型水生植物,仍以藻类居多,底部是有机质含量较少的沙质沉积物;S3和S4位于太湖东部区域,以大型水生植物为主,水环境质量较好(刘新等,2017),其中S3以挺水植物居多,S4以沉水植物为主,沉积物为含较多有机碎屑的软泥。具体点位见图 1,沉积物理化性质见表1。
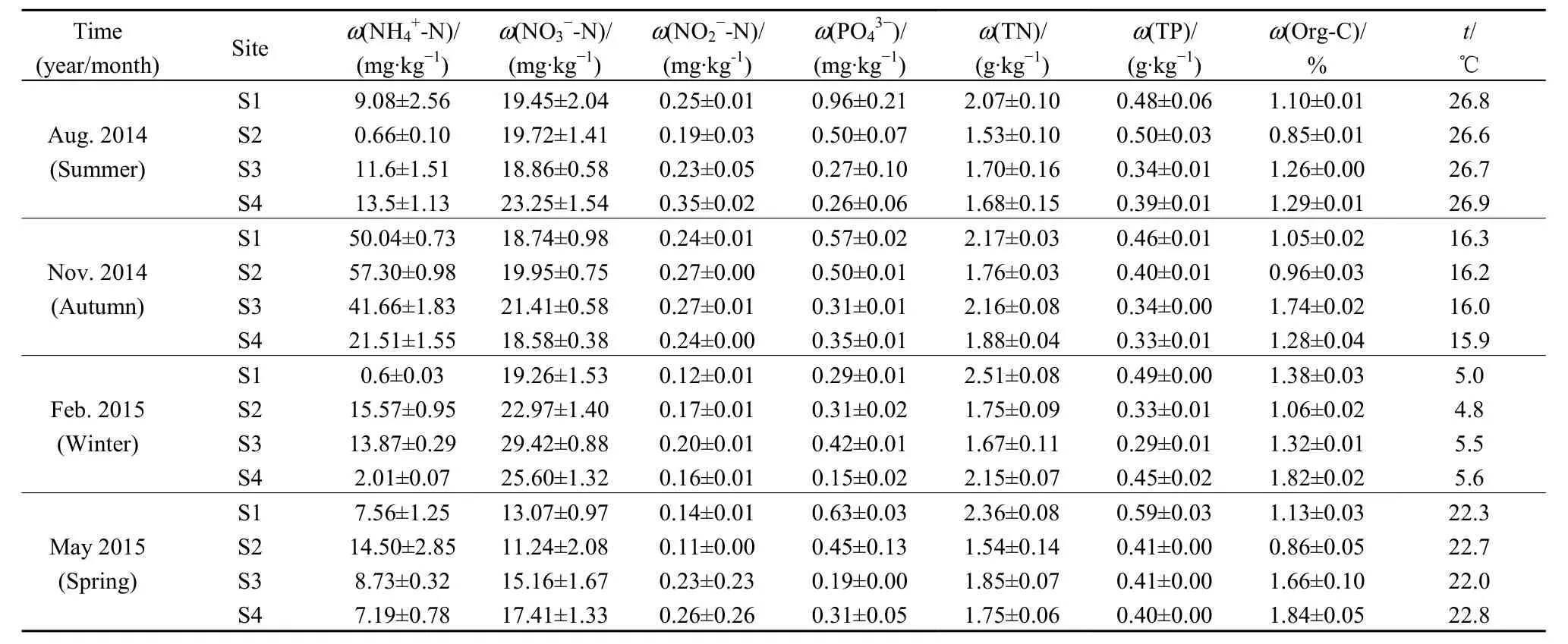
表1 沉积物理化参数(以干质量计)Table1 Physicochemicalproperties of sediment (dry mass)
1.2 沉积物样品采集
使用GPS定位,于2014年8月(夏)、2014年11月(秋)、2015年2月(冬)以及2015年5月(春)分别在点位S1、S2、S3和S4用重力采样器采集沉积物柱样3根,然后取表层2 cm沉积物放入自封袋,排出空气以保持厌氧环境,样品尽快带回实验室,分析沉积物基础化学指标,此外从各样品袋中取出部分沉积物样品供DNA提取之用。采样在每个月中旬,尽量在天气晴好、风浪较小的情况下进行,采样的同时使用多参数水质仪(YSI,USA)现场测定水温等参数。
1.3 沉积物理化性质及N2O通量测定
沉积物经冷冻干燥后过0.15 mm筛,采用碱性过硫酸钾消解-紫外分光光度法测定总氮(TN),采用钼酸盐分光光度法测定总磷(TP)(魏复盛,2002),氨氮()采用KCl浸提-蒸馏法测定,硝氮()采用酚二磺酸光度法测定,亚硝氮()采用盐酸萘乙二胺分光光度法测定,有机碳(Org-C)采用重铬酸钾氧化法测定,磷酸根()采用NaHCO3浸提-钼酸盐分光光度法测定(鲍士旦,2002)。沉积物N2O通量的测定可参照文献(钟继承等,2009)。具体操作如下:称取3 g左右的沉积物样品于50 mL血清瓶中,加入10 mL事先用高纯N2曝气过的原位湖水,然后再用高纯N2吹扫10 s,立即盖上橡胶塞以维持厌氧环境。每个样品设置两个处理,第一组处理为对照,用来采集0时刻气体样品,第二组处理振荡(70 r·min-1)避光培养4 h,用来测定沉积物N2O的通量,每个处理3个重复。采集气体样品前,剧烈摇晃血清瓶,以使顶空气体达到平衡,然后注射18 mL 3 mol·L-1NaCl溶液于血清瓶中,同时用另一支注射器收集置换出的气体,然后注入事先抽真空的气体样品瓶中。N2O采用带电子捕获器的气相色谱仪(Agilent 7890B)测定。沉积物N2O通量以单位时间内干质量沉积物的N2O生成量计。
1.4 沉积物DNA提取
沉积物DNA采用FastDNA SPIN Kit for Soil(MP,Biomedicals,USA)试剂盒提取,参照试剂盒内存放的DNA提取操作步骤,进行DNA提取和操作。用 SmartSpecTMPlus Spectrophotometer(BIO-RAD,USA)测定DNA的纯度及浓度,将提取好的DNA稀释10倍后保存于-20 ℃冰箱,原样保存于超低温冰箱备用。

表2 qPCR扩增引物及反应Table2 Amplification primer and reaction programs of qPCR
1.5 反硝化功能基因测定
反硝化功能基因nirK、nirS、norB、nosZ的丰度采用实时荧光定量PCR法(qPCR)测定,采用SYBR green染料法,在CFX96 Real-Time System(BIO-RAD,USA)仪器上运行检测。选取环境样品中得到的nirK、nirS、norB、nosZ的重组质粒,根据已知质粒浓度和阿伏伽德罗常数(6.02×1023),配置标准曲线模板,最终得到的扩增效率分别为115.2%(R2=99.5%)、86.8%(R2=99.1%)、100.6%(R2=99.5%)、101.7%(R2=99.4%),符合实验要求。qPCR反应采用20 μL体系,包括10 μL 2×SYBR premix Ex TaqTM(TaKaRa Shuzo,Shiga,Japan),20 μmol·L-1上下游引物、2 μL 模板 DNA(6-10 ng)以及7.6 μL灭菌超纯水。每个样品设置3个重复和1个阴性对照(以灭菌超纯水代替DNA作为反应模板),其qPCR扩增引物及反应程序见表2。
1.6 数据分析与处理
qPCR数据采用 CFX ManagerTM软件进行分析,沉积物理化性质、沉积物N2O通量与反硝化功能基因丰度的 Pearson相关性分析采用 SPSS 19.0处理,Origin 8.5制图。
2 结果与分析
2.1 沉积物氮磷及有机碳
各样点沉积物基础理化指标如表1所示。沉积物TN和TP全年变幅较小,其中位于梅梁弯的S1点位是典型的藻型湖区,富营化程度最高,TN明显高于其他点位,其次是位于草型湖区的S3和S4;TP是S1点位最高,其次是位于敞水区的S2,草型湖区的S3和S4最低。沉积物无机氮仅占TN含量的0.8%-4.42%。秋季沉积物氨氮含量最高,均值为(42.63±15.46) mg·kg-1,这是由于秋季水生植物和藻类逐渐死亡沉积于沉积物表层,分解矿化释放出氨,加之利用率低,进而导致氨氮升高(朱梦圆等,2011)。硝态氮是本研究湖区无机氮的主要形态,其 夏 季 均 值 为 (20.32±1.98) mg·kg-1, 与 秋 季[(19.67±1.31) mg·kg-1]无明显差异。冬季硝态氮含量最高[(24.31±4.29) mg·kg-1],春季最低[(14.22±2.66)mg·kg-1]。亚硝态氮在沉积物中无积累,均值为(0.21±0.06) mg·kg-1。PO43-含 量 较 低 , 均 值 为(0.41±0.19) mg·kg-1。沉积物有机碳存在明显的季节变化规律,整体表现为夏秋季节低、冬春季节高的趋势。大型水生植物与藻类相比有更高的生物量,更有利于所在区域有机碳的积累,故S3和S4有机碳含量明显高于S1和S2。
2.2 沉积物N2O通量
沉积物N2O通量季节变化如图2所示。各样点沉积物N2O通量季节变化规律基本一致,夏秋沉积物 N2O整体表现为负排放,其通量为-0.12-0.04 nmol·g-1·h-1,均值为-0.05 nmol·g-1·h-1。夏秋季沉积物的厌氧环境使对厌氧高敏感的Nos基因合成酶的活性显著增强并超过其他3种,有利于反硝化的彻底进行并最终生成N2(Souza et al.,2017),反硝化消耗N2O而导致N2O产生负排放。冬春季整体表现为排放,通量为-0.05-0.48 nmol·g-1·h-1,均值为 0.27 nmol·g-1·h-1。各样点沉积物 N2O 通量均在冬季达到全年最高,冬季沉积物氨氧化细菌(AOB)丰度高于夏秋季(程建华等,2016),冬季较高的有机碳含量有利于异养硝化细菌进行异氧硝化而生成N2O,而且AOB的反硝化也能产生N2O(Peng et al.,2015),这都可导致冬季沉积物产生较高的N2O通量。
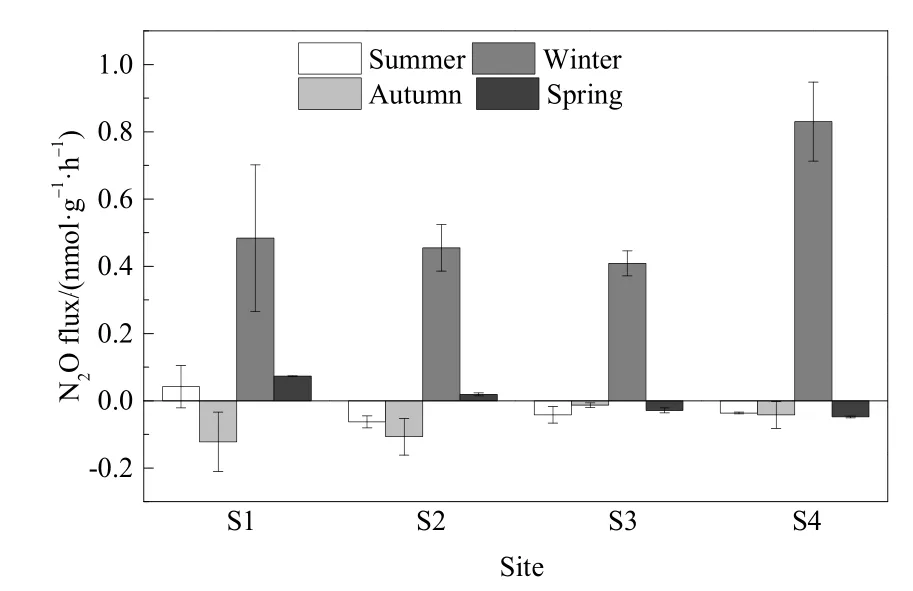
图2 沉积物N2O通量季节变化Fig.2 Seasonal variation of sediment N2O flux
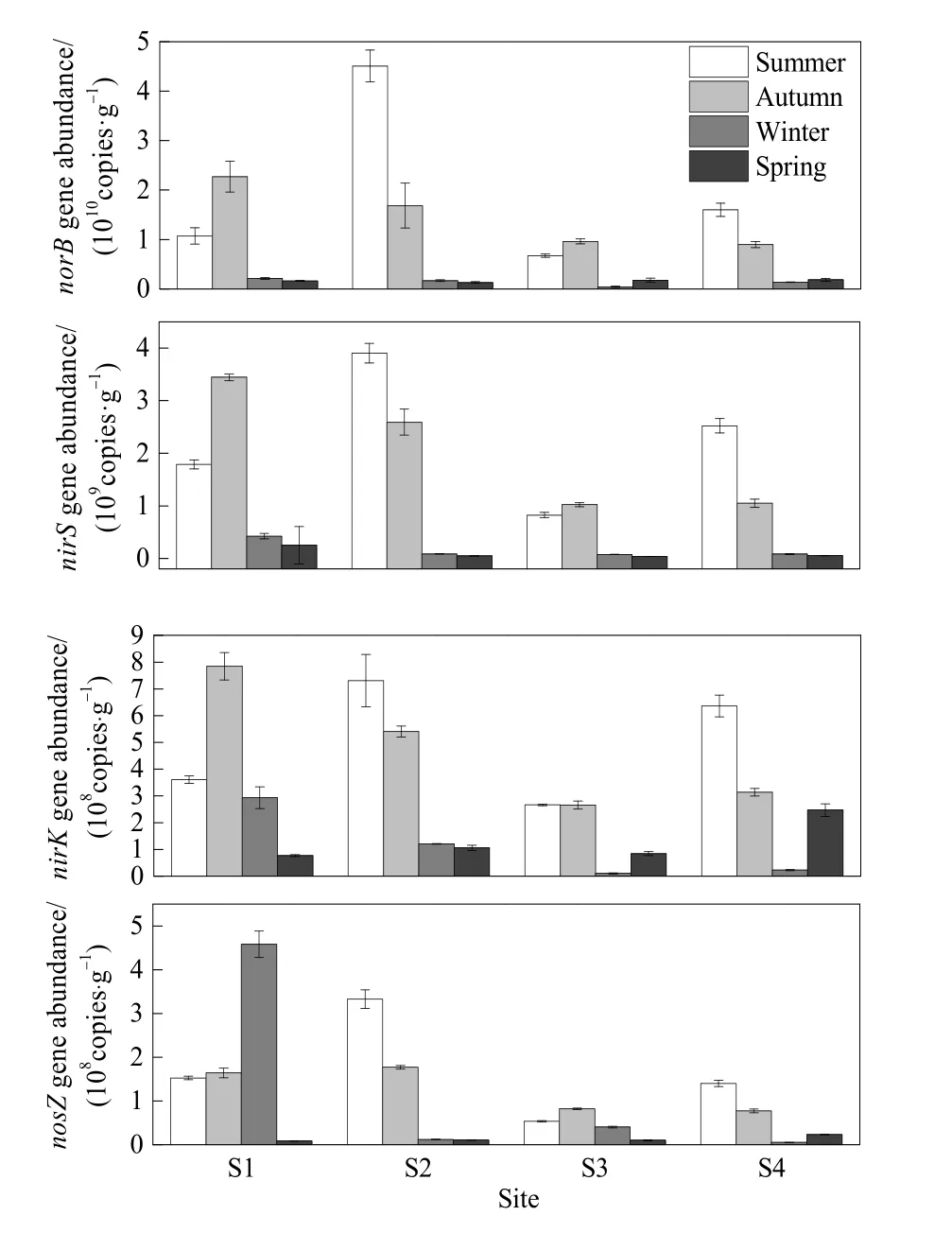
图3 沉积物反硝化功能基因丰度Fig.3 Denitrification function gene abundance in sediment
2.3 沉积物反硝化功能基因丰度
沉积物反硝化功能基因丰度季节变化如图3所示,各功能基因均大致表现为夏秋季节高、冬春季节低的特点,各样点夏秋季节差异较大,而冬春季节差异较小,与太湖西部区域沉积物nirS基因丰度季节变化(罗跃辉等,2017)规律相同。从年均值来看,各样点沉积物中 norB基因丰度最高,为9.03×109copies·g-1,其次为 nirS 基因(1.14×109copies·g-1),nirK 和 nosZ基因丰度均值分别为3.04×108copies·g-1和 1.09×108copies·g-1,S1 和 S2反硝化功能基因丰度整体大于S3和S4。
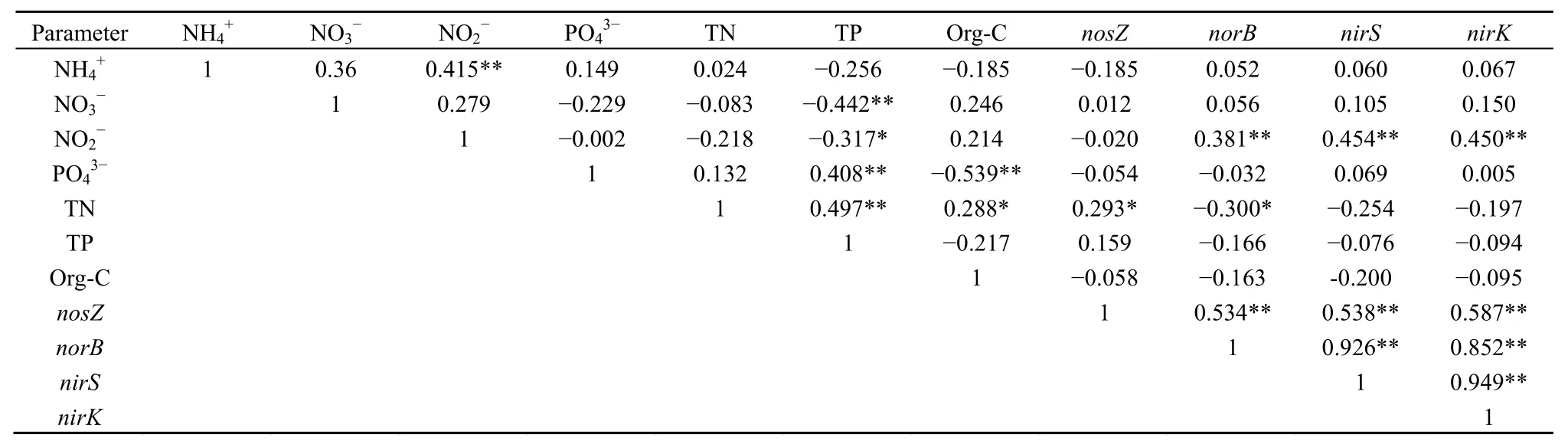
表3 反硝化功能基因丰度与沉积物化学性质Pearson相关性Table3 Pearson correlation analysis between denitrification function gene abundance and chemical properties of sediment
3 讨论
3.1 沉积物中反硝化功能基因丰度变化及其对环境因子的响应
环境因子的变化可影响微生物的数量和群落结构(Bostr et al.,1989)。沉积物理化性质与反硝化功能基因相关性分析结果如表 3所示,除 nosZ外,其余各反硝化功能基因均与沉积物含量呈极显著正相关,沉积物 TN与 nosZ呈显著正相关(P<0.05),与 norB 呈显著负相关(P<0.05),表明这两种沉积物环境因子可以用来作为影响部分反硝化功能基因丰度的指示因子,这与孙许超等(2018)的部分研究结果一致。与奉化江底泥(顾卿等,2016)和蔬菜地(王军等,2016)反硝化功能基因丰度相比,本研究太湖沉积物中,nosZ基因丰度较低,原因可能在于总氮水平较低,但也有研究发现,反硝化功能基因丰度与总氮呈负相关(高嵩涓等,2017)。在本研究中,尽管反硝化功能基因丰度与亚硝态氮关系密切,但太湖沉积物中亚硝态氮几乎无积累,且无明显四季波动(表1),故其对nirS和nirK基因丰度影响有限;与铜陵市河流沉积物中 nirS和 nirK基因丰度相当,季节变化部分一致(程建华等,2016),但与长江湿地(Hou et al.,2014)和美国河口(Lisa et al.,2014)沉积物反硝化细菌丰度的季节变化不一致,由于沉积物及土壤中的反硝化种群具有很高的多样性,且组成差异很大(Grntzig et al.,2001;Henry et al.,2004),同时受 pH(Cavigelli et al.,2000)、季节变化(Mishra et al.,2012)、土地利用方式(Rich et al.,2003)等因素影响,因而导致不同环境条件下反硝化微生物群落结构及丰度差异很大。此外,本研究中各反硝化功能基因丰度间均呈显著正相关(P<0.01),表明它们对外界环境因子的变化有基本一致的响应。
3.2 沉积物N2O通量与反硝化功能基因丰度的关系
反硝化过程可以是生成N2O的源,同时也可能是消耗N2O的汇,是这两个过程平衡的结果(Bange et al.,1996)。亚硝酸盐还原酶(nirK和 nirS)是反硝化过程中的关键酶,主导亚硝酸盐转化为氧化氮这一步反应,也是反硝化脱氮过程的限速步骤。研究发现,亚硝酸盐的积累可直接导致 N2O的排放,N2O排放量随亚硝酸盐氮的增加而增大(张静蓉等,2010),其原因在于高浓度的亚硝酸盐抑制了N2O还原酶的活性导致N2O的释放(Itokawa et al.,2001;Alinsafi et al.,2008)。本研究中夏秋季节nirK和nirS基因丰度高于冬春季,故夏秋季亚硝酸盐的消耗量大于秋冬季,从而降低了N2O的生成量,这可能是太湖沉积物N2O释放表现为夏秋季较低、冬春季较高的主要原因。相关性分析发现(表4),夏秋季沉积物nirS和nirK基因丰度与N2O通量呈极显著负相关(P<0.01),也符合上述推断。夏秋季沉积物反硝化功能基因丰度高且与 N2O通量呈极显著负相关(P<0.01),这在一定程度也表明反硝化过程进行彻底,整体表现为对N2O的消耗而产生负排放,而且N2O被同化还原为NH4+也会被消耗,导致N2O的负排放(Vieten et al.,2008)。此外,当土壤或沉积物的pH值在6-8,nosZ基因表达的酶活性不受抑制,产物主要是 N2(冯洁等,2014),太湖夏秋季沉积物间隙水pH值范围在6.7-7.2(王福芳等,2012),有利于反硝化彻底进行,这也是太湖沉积物 N2O在夏秋季表现为负排放的原因。冬春季沉积物N2O整体表现为排放(图2),而反硝化功能基因丰度明显低于夏秋季(图3),且N2O通量与各反硝化功能基因均不具显著相关性(表4),这表明反硝化并不是沉积物N2O产生的主要过程(Liu et al.,2013),这也与 Liu et al.(2018)研究结果一致。N2O可产生于硝化、反硝化、氨化以及一些化学过程,同时也会被反硝化等过程消耗,N2O通量则取决于这些过程中N2O的生成量和消耗量的差减。尽管N2O的生成与反硝化功能微生物关系密切,但仅从此方面去解释其与N2O的通量的关系,以及对反硝化过程影响的机制仍存在局限性(郑燕等,2012)。微生物DNA中反硝化功能基因的存在可指示该类微生物的反硝化能力,但并不能认为该微生物都能进行反硝化基因的表达(Nogales et al.,2002),因此,尽管大量微生物具有反硝化功能基因,但是否表达,什么特定条件下能表达,仍有待进一步研究。然而,由于样品中RNA提取和保存相对困难,相关研究仍鲜见报道。
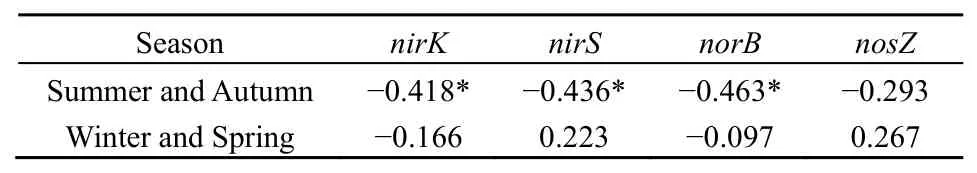
表4 沉积物N2O通量与反硝化功能基因Pearson相关性Table4 Pearson correlation analysis betweenN2O flux and denitrificationfunction gene abundance
4 结论
太湖沉积物反硝化功能基因丰度呈现夏秋季高冬春季低,具有明显的季节变化特征。各反硝化功能基因季节变化规律一致,表明它们对外界环境因子的变化有基本一致的响应。沉积物TN和NO2-是影响反硝化功能基因丰度的重要环境因子。反硝化作用并不是沉积物N2O产生的主要过程。
猜你喜欢
湖州师范学院学报(2022年9期)2022-11-09
华人时刊(2022年13期)2022-10-27
海洋通报(2022年2期)2022-06-30
环境工程技术学报(2022年3期)2022-06-05
海洋石油(2021年3期)2021-11-05
河北环境工程学院学报(2021年1期)2021-03-19
空间科学学报(2020年6期)2020-07-21
中亚信息(2016年3期)2016-12-01
中国环境科学(2016年3期)2016-02-08
金融经济(2015年16期)2015-08-21
