
隔药灸对克罗恩病大鼠结肠NLRP3炎症小体及IL-1b调节作用的实验研究
2019-02-22 10:06张霁吴丽洁李志元张丹智方圆杨延婷吴丹艳赵越李茜莹马晓芃
上海针灸杂志 2019年2期
张霁,吴丽洁,李志元,张丹,智方圆,杨延婷,,吴丹艳,赵越,李茜莹,马晓芃,
隔药灸对克罗恩病大鼠结肠NLRP3炎症小体及IL-1b调节作用的实验研究
张霁1,吴丽洁1,李志元2,张丹3,智方圆1,杨延婷1,3,吴丹艳1,赵越1,李茜莹1,马晓芃1,3
(1.上海中医药大学,上海 201203;2.浙江省中医院,杭州 310006;3.上海市针灸经络研究所,上海 200030)
通过观察隔药灸对克罗恩病大鼠结肠NLRP3炎症小体(NLRP3、ASC、Caspase-1)及下游炎症因子IL-1b表达的影响,探讨隔药灸治疗克罗恩病的抗炎机制。将清洁级雄性SD大鼠随机分为正常组、模型组、隔药灸组和假灸组4组。采用三硝基苯磺酸(2,4,6-trinitrobenzenesulfonic acid, TNBS)与乙醇混合溶液灌肠制备大鼠克罗恩病模型。造模成功后,隔药灸组取天枢(双)、气海穴进行隔药饼灸治疗;假灸组取穴、操作同隔药灸组,但不点燃艾炷。治疗结束后,记录各组大鼠结肠长度及CMDI评分;采用HE染色,光学显微镜下观察各组大鼠结肠组织形态结构,并应用免疫组织化学技术检测各组大鼠结肠NLRP3、ASC、Caspase-1及IL-1b的表达。与正常组比较,模型组大鼠结肠组织损伤严重,表现为裂隙样溃疡,炎症细胞浸润伴水肿,部分大鼠结肠黏膜可见肉芽组织形成,且结肠NLRP3、ASC、Caspase-1、IL-1b蛋白表达显著增加(均<0.05);与模型组比较,隔药灸组大鼠结肠损伤有所修复,炎症反应减轻,结肠NLRP3、ASC、Caspase-1及IL-1b蛋白表达显著降低(均<0.05);假灸组大鼠结肠损伤程度、炎症反应与模型组类似,两组结肠NLRP3、ASC、Caspase-1、IL-1b表达比较,差异均无统计学意义(>0.05)。隔药灸能下调克罗恩病大鼠结肠NLRP3、ASC、Caspase-1及IL-1b的蛋白表达,促进结肠炎性损伤的修复。
药饼灸疗法;克罗恩病;NLRP3炎症小体;间接灸;大鼠
克罗恩病(Crohn’s disease, CD)是一种慢性胃肠道炎性肉芽肿性疾病,可发生于消化道从口腔至肛门的任何部位[1]。临床症状主要包括腹泻、腹痛、体重减轻、恶心、呕吐等,在某些情况下可出现发烧或寒战,多达1/3的患者患有肛周疾病[2]。此外,研究发现多种肠外表现与CD相关,如炎症性关节炎、葡萄膜炎和虹膜炎、坏疽性脓皮病以及结节性红斑等[3]。
CD的病因及发病机制尚不明确,目前认为其发病为遗传、免疫和环境因素等交互作用的结果[3]。相关研究发现,NLR family pyrin domain containing 3 (NLRP3)基因编码的NLRP3/cryopyrin蛋白是NLRP3炎症小体的一部分,可与包含CARD结构域的细胞凋亡相关斑点样接头蛋白ASC(Apotosis-associated speck-like protein containing a CARD, ASC)结合并招募半胱天冬酶(Caspase-1)使其激活,作为调控IL-1b、IL-18生成和分泌的平台,参与包括CD在内的多种炎症性疾病的发病[4]。本课题组前期研究发现,隔药灸能下调CD大鼠结肠IL-1b的表达,减轻肠道炎症,改善结肠组织形态结构[4-5]。隔药灸是否通过调节NLRP3炎症小体下调IL-1b的表达值得进一步研究。因此,本研究拟建立大鼠CD模型,观察隔药灸对CD大鼠结肠NLRP3炎症小体(NLRP3、ASC、Caspase-1)及下游炎症因子IL-1b的调节作用,基于NLRP3炎症小体探讨隔药灸治疗克罗恩病的作用机制。
1 材料与方法
1.1 实验动物
清洁级雄性Sprague-Dawley(SD)大鼠28只,体质量为(150±20)g,由上海斯莱克实验动物有限责任公司提供[SCXK(沪)2012-0002]。饲养于上海中医药大学实验动物中心,温度为18~26℃,相对湿度为40%~70%,照明12 h昼夜交替,通风换气8~12次/h。饲料和饮用水经高压蒸汽灭菌处理,大鼠自由摄食、饮水,定期更换垫料。适应性饲养3 d,观察其饮食、活动、体态、背毛等是否健康正常,大鼠无异常后开始实验。
1.2 主要试剂和仪器
5%(w/v)2,4,6-三硝基苯磺酸(2,4,6-Trinitro- benzenesulfonic-acid, TNBS)(Sigma公司,美国),无水乙醇、戊巴比妥钠(国药集团化学试剂有限公司);NLRP3一抗(Abcam公司,英国),ASC一抗(Abcam公司,英国),Caspase-1一抗(Abcam公司,英国),IL- 1b一抗(Abcam公司,英国);Dako REAL EnVision兔鼠通用二抗检测试剂盒(Dako,丹麦),组织脱水机(Leica公司,德国),组织包埋机(Leica公司,德国),石蜡切片机(Leica公司,德国),石蜡切片水浴缸(Leica公司,德国),烤片台(Leica公司,德国)。
1.3 模型制备
参照Morris方法[6],将5%(w/v)的TNBS与50%乙醇按照体积比2:1混合成灌肠液。大鼠称重后,用超纯水配制1%戊巴比妥钠溶液,按照30 mg/kg剂量腹腔注射麻醉。造模开始前24 h禁食不禁水,使用大鼠灌胃器自肛门插入肠道6~8 cm,以3 mL/kg的剂量灌入上述灌肠液,灌肠后保持大鼠倒立位60 s,每7 d进行1次,持续4周。于造模结束时,每组随机取1只大鼠处死,取其结肠组织做HE染色以检验模型成功与否。在验证模型制备成功的基础上,进行干预。
1.4 分组与治疗
正常组不进行任何处理与治疗。
模型组在模型制备成功后,不进行任何治疗,只作与隔药灸组相同的抓取和固定。
隔药灸组选取天枢(双)、气海穴,进行隔药灸治疗,每次每穴各灸2壮(约10 min)。每日1次,共治疗7次。药饼主要成份为附子粉,灸前用黄酒调和,并用动物专用模具制成药饼;用模具将艾绒压实制成锥形艾炷,艾炷重量为90 mg,置于药饼上,点燃艾炷施灸。
假灸组仅做与隔药灸组相同的固定,并将艾炷及药饼置于大鼠腹部天枢(双)、气海穴上,但不点燃艾炷,每次10 min。每日1次,共7次。
1.5 样本采集与处理
治疗结束后,各组大鼠禁食、不禁水24 h,用1%戊巴比妥钠,按照30 mg/kg剂量腹腔注射麻醉,腹主动脉取血。沿大鼠腹正中线剪开腹部皮肤,充分暴露直肠、结肠部分,自耻骨联合处至盲肠取下大鼠整个结肠,并测量结肠长度并记录数据后,沿肠系膜纵轴剖开,取肛门起向上5~8 cm部分,用4℃冷生理盐水冲洗,取下结肠段放入4%多聚甲醛溶液内固定24 h。
1.6 观察指标
1.6.1 结肠长度
测量各大鼠结肠长度,并进行统计分析。
1.6.2 结肠大体形态损伤指数(colon macroscopic damage index, CMDI)
对各组大鼠结肠进行结肠大体损伤评分,该评分包括结肠及周围组织的黏连程度、溃疡及炎症损伤的严重程度。无黏连为0分;轻度黏连(结肠与其他组织剥离较易)为1分;重度黏连为2分。无溃疡及炎症为0分;局部充血无溃疡为1分;1处溃疡不伴出血和肠壁增厚为2分;1处溃疡伴炎症为3分;2处溃疡伴炎症为4分;>2处溃疡伴炎症或炎症面积>1 cm为5分;溃疡和(或)炎症面积2 cm随病变范围每增加1 cm,积分增加1分(6~8分)。根据以上评分标准进行评分,并分析统计[7]。
1.6.3 结肠组织形态学观察
采用苏木素-伊红(hematoxylin-eosin, HE)对组织切片进行染色,光学显微镜下观察各组大鼠结肠组织形态学变化。HE染色主要步骤如下,①常规脱蜡水化,切片放入二甲苯Ⅰ、Ⅱ中脱蜡各20 min,梯度乙醇100%、90%、80%、70%各3 min。②双蒸水冲洗。③核染色,用苏木素染色约2.5 min后,放入自来水中缓缓冲洗10 min;1%盐酸乙醇分化2 s,再流水缓缓冲洗 5 min反蓝。④伊红胞质染色,将切片放入0.1%伊红中染色10 min后,依次经过梯度乙醇70%、80%各1 min, 90%、100%各3 min,根据镜下显色调整时间。⑤透明,将切片放入二甲苯Ⅰ、Ⅱ各15 min后,擦去多余液体,中性树胶封片。
1.6.4 结肠组织NLRP3、ASC、Caspase-1及IL-1b蛋白表达检测
采用免疫组织化学Envision法检测。主要步骤如下,①结肠组织石蜡切片(4mm)于60℃恒温烤箱中烤片1.5 h。②脱蜡水化,将切片浸于二甲苯Ⅰ、Ⅱ中各10 min,梯度乙醇100%、90%、80%、70%各3 min,PBS冲洗3 min×3次。③抗原修复,将切片置于600 mL柠檬酸盐缓冲液中,微波中高火沸腾后,取出室温冷却10 min,再沸腾8 s,于室温自然冷却1 h。④消除内源性过氧化物酶活性,将组织切片浸于3%的H2O2中,避光孵育25 min后,双蒸水冲洗3 min×3次。⑤玻片擦干,用5%BSA封闭液封闭,室温孵育20 min,甩去多余液体,滴加一抗,4℃冰箱过夜。37℃恒温烤箱复温45 min, PBS冲洗3 min×3次。滴加A液,室温孵育30 min后,PBS冲洗3 min×3次。擦干,滴加DAB工作液(10mL C液:990mL B液),光镜下控制显色,显色完全后,湿盒中止显色1 min。苏木素染色2.5 min,自来水冲洗10 min。⑥1%盐酸乙醇分化1~3 s,自来水冲洗5 min反蓝。⑦梯度乙醇脱水70%、80%、90%、100%各3 min。⑧二甲苯Ⅰ、Ⅱ各15 min透明。⑨封片(中性树胶:二甲苯=1:1.5)。⑩数据采集,每张切片随机选取5个视野拍摄照片,通过Image-pro plus 6.0软件进行半定量分析,取阳性目标的积分光密度进行统计。
1.7 统计学方法
采用SPSS18.0软件进行数据统计。对所有计量资料进行正态分布检验和方差齐性检验。当计量资料符合正态分布时采用均数±标准差表示;当计量资料呈偏态分布时采用中位数(最小值,最大值)的形式表示。若计量资料服从正态分布,且数据符合方差齐性检验,组间差异采用单因素方差分析(),两两比较用最小显著差法(,),方差不齐时用比较组间差异。若计量资料不服从正态分布,则用秩和检验。以<0.05表示差异有统计学意义。
2 结果
2.1 大鼠结肠长度的变化
与正常组比较,模型组、隔药灸组、假灸组大鼠结肠长度均缩短,差异均有统计学意义(<0.05)。与模型组比较,隔药灸组大鼠结肠长度虽有增加趋势,但两组比较差异无统计学意义(>0.05)。而假灸组大鼠结肠长度与模型组比较差异无统计学意义(>0.05)。详见表1。
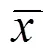
表1 各组大鼠结肠长度比较 (±s,cm)
注:与正常组比较1)<0.05
2.2 大鼠CMDI的变化(表2)
表2 各组大鼠CMDI比较 (±s,分)
注:与正常组比较1)<0.05;与模型组比较2)<0.01
与正常组比较,模型组、假灸组大鼠CMDI升高,差异均有统计学意义(<0.05)。与模型组比较,隔药灸组大鼠CMDI明显降低,两组比较差异有统计学意义(<0.01),假灸组大鼠CMDI与模型组比较差异无统计学意义(>0.05)。详见表2。
2.3 大鼠结肠组织形态学变化
光学显微镜下观察,正常组大鼠结肠黏膜层上皮细胞形态结构完整,固有层杯状细胞、腺体排列整齐有序;黏膜下层疏松结缔组织形态完整;肠壁肌层(内环、外纵)结构完整无增生,浆膜层分布清晰。模型组大鼠结肠炎症反应明显,局部黏膜表面有糜烂甚至溃疡形成,溃疡呈裂隙状,局部杯状细胞增生明显,腺体破坏、腺腔排列紊乱;黏膜下层结缔组织严重水肿伴炎症细胞浸润,大量纤维组织增生;肠壁肌层、浆膜层分布尚清晰,病理变化较接近克罗恩病。隔药灸组大鼠结肠黏膜层结构尚完整,损伤处可见上皮组织修复,杯状细胞及腺体修复较好,腺体排列尚规则;黏膜固有层及黏膜下层可见炎症细胞浸润,黏膜下层结缔组织中度水肿;结肠肌层、浆膜层形态结构完整。假灸组大鼠结肠组织病理学表现与模型组类似。结果提示,隔药灸对克罗恩模型大鼠结肠炎症损伤具有一定的改善作用。详见图1。
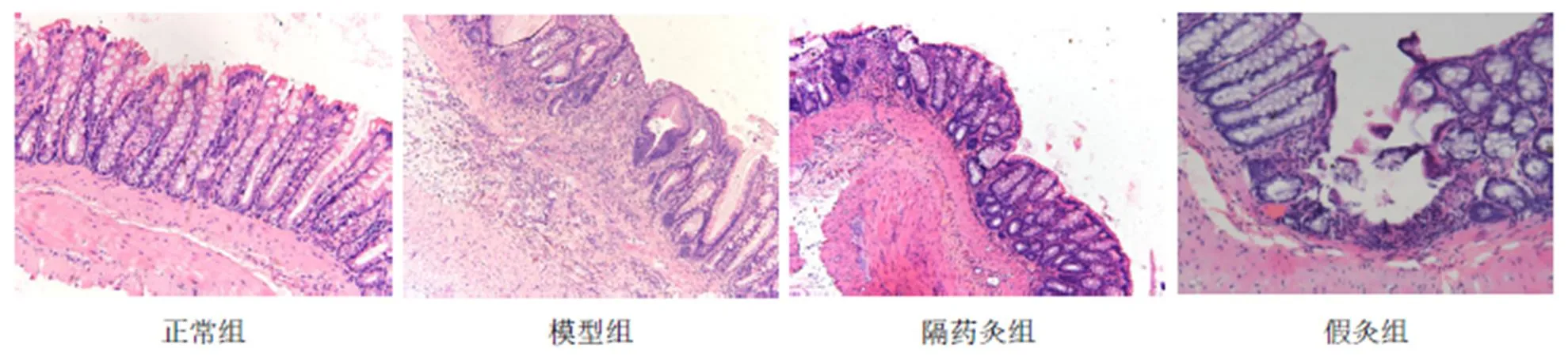
图1 各组大鼠结肠组织HE染色结果(×200)
2.4 大鼠结肠NLRP3蛋白表达的变化(表3)
表3 各组结肠NLRP3蛋白表达结果比较 (±s)
注:与正常组比较1)<0.01;与模型组比较2)<0.01; 与假灸组比较3)<0.05
与正常组比较,模型组、假灸组大鼠结肠NLRP3蛋白表达明显增高(均<0.01);与模型组比较,隔药灸组大鼠结肠NLRP3蛋白表达明显减少(<0.01),而假灸组大鼠结肠NLRP3蛋白表达则无显著性变化 (>0.05)。与假灸组比较,隔药灸组大鼠结肠NLRP3蛋白表达明显降低(<0.05)。详见表3、图2。
2.5 大鼠结肠ASC蛋白表达的变化
与正常组比较,模型组、假灸组大鼠结肠ASC蛋白表达明显增高(均<0.01);与模型组比较,隔药灸组大鼠结肠ASC蛋白表达显著下降(<0.01),假灸组大鼠结肠ASC表达无显著性变化(>0.05);与假灸组比较,隔药灸组大鼠结肠ASC表达明显降低(<0.01)。详见表4、图3。
表4 各组大鼠结肠ASC表达结果比较 (±s)
注:与正常组比较1)<0.01;与模型组比较2)<0.01; 与假灸组比较3)<0.01
2.6 大鼠结肠Caspase-1蛋白表达的变化
与正常组比较,模型组、假灸组大鼠结肠Caspase-1蛋白表达明显增高(均<0.05);与模型组比较,隔药灸组大鼠结肠Caspase-1蛋白表达明显降低(<0.01),假灸组大鼠结肠Caspase-1表达无显著性变化(>0.05);与假灸组比较,隔药灸组大鼠结肠Caspase-1蛋白表达明显降低(<0.01)。详见表5、图4。
表5 各组结肠Caspase-1表达结果比较 (±s)
注:与正常组比较1)<0.01;与模型组比较2)<0.01; 与假灸组比较3)<0.01
2.7 大鼠结肠IL-1b蛋白表达的变化
与正常组比较,模型组、假灸组大鼠结肠IL-1b蛋白表达明显增高(均<0.05);与模型组比较,隔药灸组大鼠结肠IL-1b蛋白表达明显降低(<0.05),假灸组大鼠结肠IL-1b蛋白表达无显著变化(>0.05);与假灸组比较,隔药灸组大鼠结肠IL-1b蛋白表达明显降低(<0.01)。详见表6、图5。
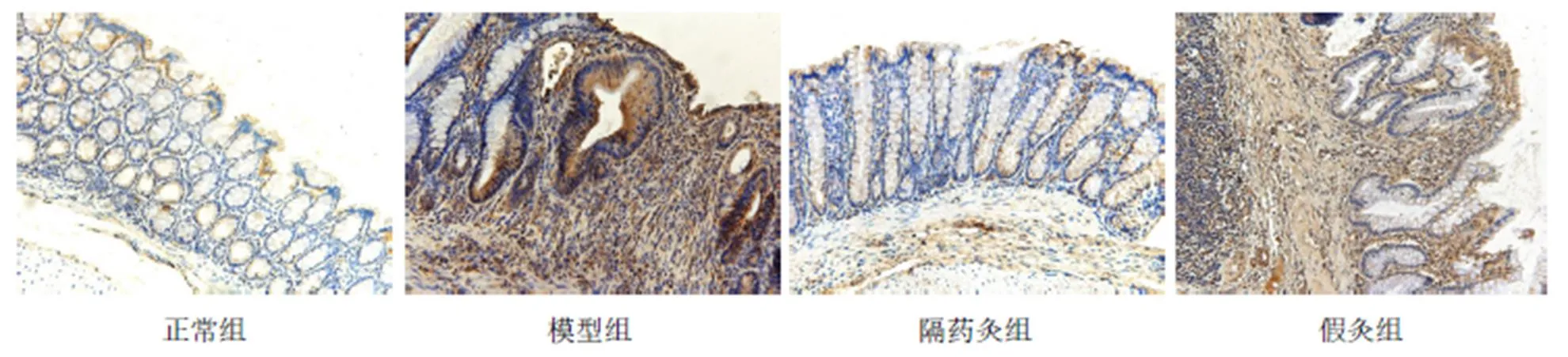
图2 各组大鼠结肠组织NLRP3的免疫组化(×200)
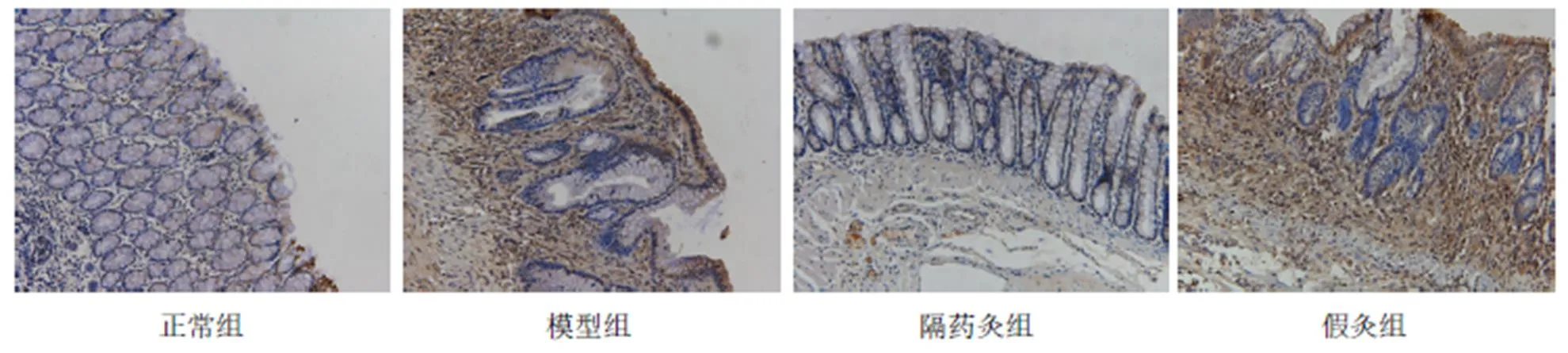
图3 各组大鼠结肠组织ASC表达的免疫组化(×200)
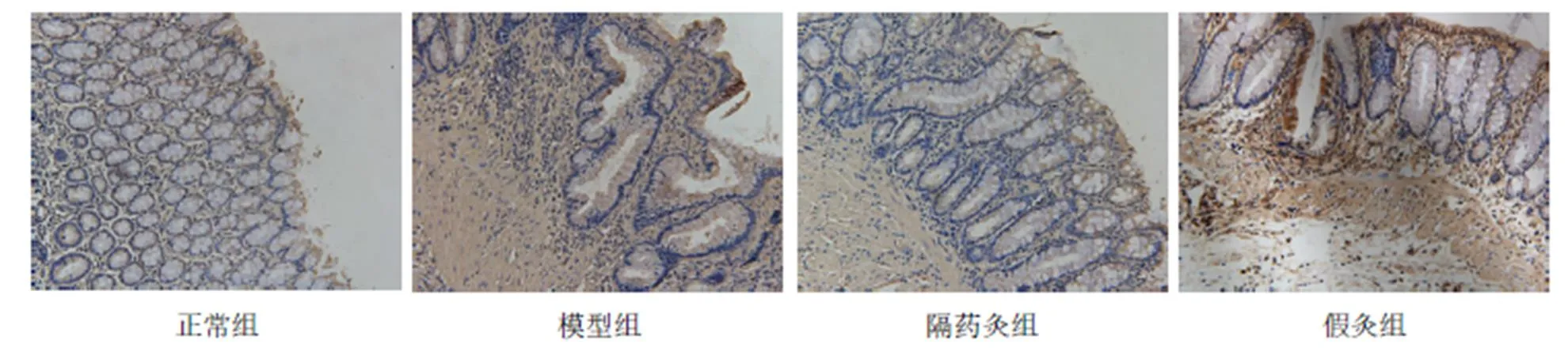
图4 各组大鼠结肠组织Caspase-1的免疫组化(×200)
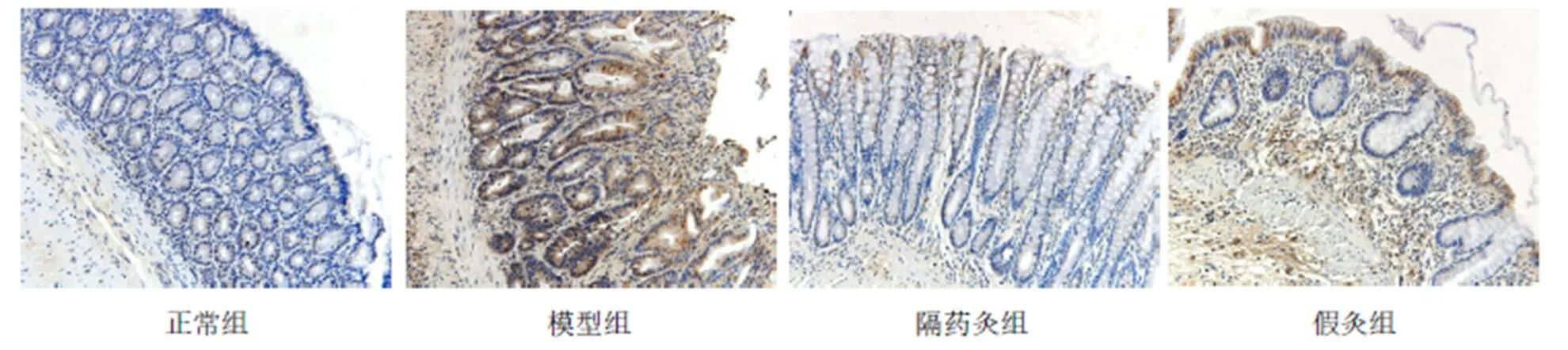
图5 各组大鼠结肠组织IL-1β的免疫组化(×200)
表6 各组大鼠结肠IL-1b表达结果比较 (±s)
注:与正常组比较1)<0.01;与模型组比较2)<0.01; 与假灸组比较3)<0.01
3 讨论
克罗恩病是一种慢性、复发性疾病,组织活检可见肉芽肿性透壁性炎症,可累及消化道任何部分,但最常见于末端回肠和邻近结肠,呈不连续性、节段性病位分布,目前临床上尚未有治愈手段,部分患者选择外科手术切除治疗[8]。重症CD患者迁延难愈,预后不良,是公认的引发结肠肿瘤的因素[9]。根据流行病学调查显示,不同种族间CD的发病率差异显著,1项基于人群的系统评价研究显示,CD发病率最高的是欧洲(德国CD患病率为322/10万),北美次之(美国CD发病率为319/10万)。其中22项关于CD的研究中有16项(72.7%)研究显示北美和欧洲CD的发病率呈稳定或下降趋势;而自1990年以来,非洲、亚洲和南美洲的新兴工业化国家(包括巴西)的CD发病率一直呈上升趋势[10];近年来我国CD的发病率逐年上升,如今已成为国内消化科的常见病[11]。由于其复发缓解过程会对不同年龄患者(尤其是年轻人)的生活质量产生显著影响,且尚未有有效治愈该疾病的药物,故有30%~50%的炎症性肠病患者选择常规治疗外的其他疗法作为缓解手段[12-13]。而且随着病程的进展,肠纤维化、炎症等会导致肠狭窄,最终会出现梗阻症状。尽管近年来对CD的研究取得了一定进展,抗炎等治疗显著改善了患者临床症状,但现有的治疗仍无法改善患者的肠纤维化甚至狭窄[14]。因此,探寻CD的有效治疗方法一直是医学界关注的热点。
研究显示,在CD发病过程中固有免疫系统发挥关键作用,肠黏膜固有免疫系统主要由肠道黏膜屏障、固有免疫细胞及其产物组成[15]。黏膜屏障一旦受损则细菌及其产物入侵黏膜下层,与初始免疫细胞和过继免疫细胞发生免疫反应,释放促炎因子(TNF-a、IL-1、IL-6、ROS等)放大炎症反应。NLRP3是一种存在于胞质内的模式识别受体,在识别特异性的病原相关分子模式后,将信号传导进入细胞,激活细胞内核转录因子NF-kB,进入细胞核后启动一系列相关炎症因子转录及炎症小体装配的级联反应。其中核苷酸寡聚结构域受体(NLRs/ALRs)被激活并招募胞质中的相关蛋白形成复合体即炎症小体(inflammasome),炎症小体的装配可激活pro-Caspases-1和Caspases-11,将酶原形式的Caspase快速转化为活性蛋白酶,切割前体pro- IL-1b、pro-IL-18为成熟的炎症因子并分泌成熟的IL-1b、IL-18[16]。NLRP3炎症小体的激活失控,则会产生过量IL-1b、IL-18或其他炎性因子,进而诱导大量炎性介质、炎性细胞等在患处堆积,导致机体发生严重炎症并诱发坏死及损伤。CD发病时,肠黏膜被肠道中各种刺激因子刺激,导致NLRP3炎症小体激活,随之IL-1b等表达增多,并造成局部黏膜损伤[17-18]。
另外,有研究发现CD的易感性与NLRP3炎症小体和NOD2的异常激活相关[19]。随着炎症小体与CD的相关性研究的深入,发现NLRP3基因编码区(先前为NALP3)和CARD8基因(编码CARD蛋白)中的单核苷酸多态性(SNPs)与CD的易感性有关[20]。一项研究对来自CD家族的710个样本组合进行分析,发现NLRP3基因调节区中的3个SNP与CD具有相关性[21]。另一项研究发现,纳入研究的25名CD患者中有60%的患者检测到NLRP3炎症小体激活,而溃疡性结肠炎患者与对照组相比较NLRP3炎症小体的激活差异无统计学意义[18]。以上研究提示,NLRP3炎症小体的激活与CD发生发展关系密切,深入研究NLRP3炎症小体与CD之间的关系,有助于进一步揭示CD的发病机制。
中医学中无CD的专属名词,依据其症状常被归类为“泄泻”“腹痛”等范畴。艾灸是中医学重要的组成部分,具有调整阴阳、扶正祛邪、疏通经络等作用。作为中医学治疗肠腑疾病的有效疗法,艾灸近年来在炎症性肠病的临床治疗中渐渐被广泛应用,对于轻、中度CD患者的疗效较为明显[22-24]。大量临床与动物实验研究结果也证实,艾灸治疗炎症性肠病可能通过调节炎症反应通路、炎性因子水平、基因表达等,起到降低炎症反应、修复结肠黏膜屏障、减少组织纤维化、减轻内脏痛、缓解焦虑情绪等作用[5,25]。
本研究结果显示,CD模型大鼠结肠组织的NLRP3、ASC、Caspase-1、IL-1b蛋白表达均显著升高。Du X等[26]通过TNBS诱导SD大鼠结肠炎模型,模型组大鼠结肠NLRP3、IL-1b表达亦显著升高。Castro J等[27]通过TNBS诱导大鼠结肠炎模型,并采用RT-PCR检测结肠NLRP3、IL-1bmRNA表达,发现两者在模型组结肠组织中的表达均显著上升。Bauer C等[28]使用DSS诱导急性结肠炎模型,发现NLRP3基因缺陷型(NLRP3-/-)小鼠与野生型小鼠相比较,结肠损伤极大程度降低; NLRP3炎症小体及IL-1b的表达显著上升,且IL-1b的表达依赖NLRP3炎症小体激活。以上研究结果与本研究结果均有一致性,提示CD模型NLRP3炎症小体及其下游炎症因子IL-1b蛋白的表达异常增高。本研究在隔药灸治疗后,CD大鼠结肠组织病理损伤显著改善,黏膜层上皮、杯状细胞及腺体得到一定程度的修复,炎症反应减轻;结肠组织中NLRP3炎症小体(NLRP3、ASC、Caspase-1)及其下游炎症因子IL-1b蛋白的表达水平降低。IL-1b是NLRP3炎症小体信号通路下游重要的炎症因子,与肠道炎症相关,已有研究证实IL-1b是引发肠道炎症最重要的致炎因子之一,可导致肠道屏障功能障碍,诱发并扩大炎症反应等[29-32]。因此,推测隔药灸可能通过抑制CD大鼠结肠NLRP3炎症小体的异常激活,进而下调结肠炎症因子IL-1b蛋白的表达,达到促进结肠炎性损伤修复之目的。
炎症小体的发现为CD发病机制以及艾灸抗炎机制的研究提供了新的视角。阐明炎症小体活化及其调控CD炎症反应的确切分子机制,对于CD的防治具有重要意义,并有望成为治疗CD的新靶点。通过研究NLRP3炎症小体上下游信号通路与CD发病的关系以及艾灸对其的调控作用,将有助于进一步提高CD的临床疗效。
[1] Abraham C, Cho JH. Inflammatory bowel disease[J]., 2009,361(21):2066-2078.
[2] Feuerstein JD, Cheifetz AS. Crohn Disease: Epidemi- ology, Diagnosis, and Management[J]., 2017,92(7):1088-1103.
[3] Vavricka SR, Schoepfer A, Scharl M,. Extra- intestinal manifestations of inflammatory bowel disease[J]., 2015,21(8):1982- 1992.
[4] Baumgart DC, Sandborn WJ. Crohn's disease[J]., 2012,380(9853):1590-2605.
[5] 魏凯,张丹,窦传字,等.艾灸对克罗恩病大鼠结肠NF-kB p65及TNF-a、IL-1b调节作用的研究[J].世界中医药,2013,8(8):862-866.
[6] Morris GP, Beck PL, Herridge MS,. Hapten-induced model of chronic inflammation and ulceration in the rat colon[J]., 1989,96(2):795-803.
[7] Paiotti AP, Miszputen SJ, Oshima CT,. Etanercept attenuates TNBS-induced experimental colitis: role of TNF-α expression[J]., 2011, 42(5):443- 450.
[8] Cheifetz AS. Management of active Crohn disease[J]., 2013,309(20):2150-2158.
[9] Zaki MH, Vogel P, Body-Malapel M,. IL-18 production downstream of the Nlrp3 inflammasome confers protection against colorectal tumor formation[J]., 2010,185(8):4912-4920.
[10] Ng SC, Shi HY, Hamidi N,. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: a systematic review of population-based studies[J]., 2018,390(10114):2769-2778.
[11] Molodecky NA, Soon IS, Rabi DM,. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review[J]., 2012,142(1):46-54.e42; quiz e30.
[12] Sirois FM. Health-related self-perceptions over time and provider-based Complementary and Alternative Medicine
(CAM) use in people with inflammatory bowel disease or arthritis[J]., 2014,22(4):701- 709.
[13] Opheim R, Bernklev T, Fagermoen MS,. Use of complementary and alternative medicine in patients with inflammatory bowel disease: results of a cross-sectional study in Norway[J]., 2012,47 (12):1436-1447.
[14] Spinelli A, Correale C, Szabo H,. Intestinal fibrosis in Crohn's disease: medical treatment or surgery?[J]., 2010,11(2):242-248.
[15] Oshima T, Miwa H. Gastrointestinal mucosal barrier function and diseases[J]., 2016,51(8): 768-778.
[16] Kayagaki N, Warming S, Lamkanfi M,. Non- canonical inflammasome activation targets caspase-11[J]., 2011,479(7371):117-121.
[17] Liu L, Dong Y, Ye M,. The pathogenic role of NLRP3 inflammasome activation in Inflammatory bowel diseases of both mice and humans[J]., 2017,11(6):737-750.
[18] Lazaridis LD, Pistiki A, Giamarellos-Bourboulis EJ,. Activation of NLRP3 inflammasome in inflammatory bowel disease: differences between Crohn's disease and ulcerative colitis[J]., 2017,62(9):2348- 2356.
[19] Elia PP, Tolentino YF, Bernardazzi C,. The role of innate immunity receptors in the pathogenesis of inflammatory bowel disease[J]., 2015: 936193.
[20] Schoultz I, Verma D, Halfvarsson J,. Combined polymorphisms in genes encoding the inflammasome components NALP3 and CARD8 confer susceptibility to Crohn's disease in Swedish men[J]., 2009,104(5):1180-1188.
[21] Villani AC, Lemire M, Fortin G,. Common variants in the NLRP3 region contribute to Crohn's disease susceptibility[J]., 2009,41(1):71-76.
[22] Joos S, Brinkhaus B, Maluche C,. Acupuncture and moxibustion in the treatment of active Crohn's disease: a randomized controlled study[J]., 2004,69(3): 131-139.
[23] 包春辉,吴璐一,吴焕淦,等.针灸治疗活动期克罗恩病:随机对照研究[J].中国针灸,2016,36(7):683-688.
[24] Shang HX, Wang AQ, Bao CH,. Moxibustion combined with acupuncture increases tight junction protein expression in Crohn's disease patients[J]., 2015,21(16):4986-4996.
[25] Wei K, Zhang D, Hong J,. Herb-partitioned moxibustion and the miRNAs related to Crohn's disease: a study based on rat models[J]., 2015:265238.
[26] Du X, Chen W, Wang Y,. Therapeutic efficacy of carboxyamidotriazole on 2,4,6-trinitrobenzene sulfonic acid-induced colitis model is associated with the inhibition of NLRP3 inflammasome and NF-kappaB activation[J]., 2017,45:16-25.
[27] Castro J, Ocampo Y, Franco L. Cape Gooseberry [Physalis peruviana L.] Calyces Ameliorate TNBS Acid- induced Colitis in Rats[J]., 2015,9(11): 1004-1015.
[28] Bauer C, Duewell P, Mayer C,. Colitis induced in mice with dextran sulfate sodium (DSS) is mediated by the NLRP3 inflammasome[J]., 2010,59(9):1192- 1199.
[29] De Santis S, Kunde D, Galleggiante V,. TNFalpha deficiency results in increased IL-1beta in an early onset of spontaneous murine colitis[J]., 2017, 8(8):e2993.
[30] Ogawa M, Osada H, Hasegawa A,. Effect of interleukin-1beta on occludin mRNA expression in the duodenal and colonic mucosa of dogs with inflammatory bowel disease[J]., 2018,32(3):1019- 1025.
[31] 张璐,王诗怡,范一宏,等.克罗恩病患者小肠细菌过生长的监测及意义研究[J].浙江临床医学,2017,19(6): 1029-1032.
[32] 吴璐一,翁志军,吴焕淦,等.Th17细胞因子在CD大鼠结肠不同炎性时期的差异表达[J].世界华人消化杂志,2016,24(14):2161-2169.
Experimental Study on the Regulating Effect of Herbal Cake-partitioned Moxibustion on NLRP3 Inflammasome and IL-1β in Colons of Crohn's Disease Rats
1,1,2,3,1,1,3,1,1,1,1,3.
1.,201203,; 2.,310006,;3.,200030,
To explore the action mechanism of moxibustion in treating Crohn's disease (CD) by observing the effect of herbal cake-partitioned moxibustion on the expression of NLRP3 inflammasome (NLRP3, ASC and Caspase-1) and IL-1β.Male Sprague Dawley (SD) rats of clean conventional grade were randomized into a normal control group (NG), a model control group (MG), a herbal cake-partitioned moxibustion control group (MoxG), and a sham moxibustion control group (SMG). The CD rat models were developed by using the mixture of 2,4,6-trinitrobenzene sulfonic acid (TNBS) and alcohol via enema. When the models were successfully developed, the MoxG received moxibustion intervention at bilateral Tianshu (ST25) and Qihai (CV6), and the SMG were administered by unlighted moxa cones. At the end of treatment, the length of each rats' colon and the colon macroscopic damage index (CMDI) score were recorded, the histopathological variations of rats' colons were observed by adopting HE staining and light microscope, and the expressions of NLRP3, ASC, Caspase-1 and IL-1β in rats' colons were determined by using immunohistochemical technique.Compared to the NG, the MG had its rats' colons present with severe damages, fissured ulcers and inflammatory cell infiltration with edema, and granulomas in submucosa of some colons, and its expressions of NLRP3, ASC, Caspase-1 and IL-1β increased significantly (<0.05); compared to the MG, the MoxG had rats' colons present with improved structures and reduced intestinal inflammation, and its expressions of NLRP3, ASC, Caspase-1 and IL-1β dropped significantly (<0.05); the SMG had its rats' colon inflammation present similarly to the MG, and its expressions of NLRP3, ASC, Caspase-1 and IL-1β had no significant difference (>0.05).Herbal Cake-partitioned moxibustion can down-regulate the expressions of NLRP3, ASC, caspase-1 and IL-1β in CD rats' colons to promote the repair of colon damage.
Medicinal cake-partitioned moxibustion; Crohn’s disease; NLRP3 inflammasome; Indirect moxibustion; Rats
1005-0957(2019)02-0119-08
R2-03
A
10.13460/j.issn.1005-0957.2019.02.0119
2018-08-03
国家重点基础研究发展计划(2015CB554501);国家自然科学基金项目(81674073);上海市卫生计生系统优秀学科带头人培养计划项目(2017BR047)
张霁(1990—),女,2016级博士生,Email:zhangdaimao1990@163.com
吴丽洁(1990—),女,2017级博士生,Email:wljzwb@163.com
马晓芃(1973—),女,研究员,博士生导师,Email:pengpengma@163.com
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
昆明医科大学学报(2021年8期)2021-08-13
中国民间疗法(2020年22期)2021-01-07
医学新知(2019年4期)2020-01-02
中国内镜杂志(2017年2期)2017-03-20
中国内镜杂志(2017年2期)2017-03-20
腹腔镜外科杂志(2016年10期)2016-06-01
中国医疗美容(2015年1期)2015-07-12
中国卫生标准管理(2015年3期)2015-01-27
中国卫生标准管理(2015年13期)2015-01-26
