
白令海鱼类分类多样性及鱼类区系研究❋
2019-02-21 09:23林龙山宋普庆陈永俊
中国海洋大学学报(自然科学版) 2019年4期
张 然, 林龙山, 李 渊, 宋普庆, 陈永俊, 张 静
(1.国家海洋局第三海洋研究所,福建 厦门 361005; 2.厦门大学海洋与地球学院 福建 厦门 361102; 3.集美大学水产学院,福建 厦门 361012)
白令海位于太平洋最北端,由东北部的浅海大陆架和西南部的深水盆地组成,通过白令海峡与北冰洋相连,是全球气候变化和反馈最敏感的地区之一[1-3]。白令海具有较高的海洋生态系统生产力和丰富的渔业资源,气候变暖引起北极地区海水表面温度上升、冻土层融化、海冰覆盖面积减少等对白令海物种组成产生了深刻影响,其中鱼类作为海洋食物链中的重要一环,对环境变化较为敏感,受到国内外学者广泛关注[4-8]。
鱼类物种组成是不同鱼类种群之间相互联系及其与所处环境条件综合因子的长期影响和适应过程中逐渐形成的[9-10]。白令海地处北太平洋中高纬度海域,北部陆架区存在着强大的夏季冷水团,受到海冰季节性消融的影响[11],南部经由阿留申岛链上的诸水道与北太平洋水交换顺畅,水体性质深受北太平洋入流水的影响,底层水及中层水物理性质与北太平洋水体相近[12],因此形成了白令海独特的水文特征及鱼类物种组成。本研究根据Fish Base[13]、Global Species[14]等数据库中的鱼类物种资料,并结合中国第四次北极科学考察在白令海的鱼类调查数据,从分类多样性的角度对该海域的鱼类进行了分析。分类多样性分析是从物种间的亲缘关系及分类水平上对某一区域的物种进行的多样性评估[15-17],对于鱼类分类多样性的研究,国内外均有报道[18-20],国内主要集中于我国近海,如黄海、大亚湾、北部湾及南海珊瑚礁等[21-24],北极海域未见报道。本研究初次分析了白令海鱼类分类多样性,并将该海域与楚科奇海、北太平洋边缘海、黄海、南海等不同纬度海域的鱼类多样性水平进行比较,探讨了白令海鱼类多样性水平与地理环境之间的关系,以期对白令海的鱼类组成有更加全面地了解,为进一步探究北极海域太平洋扇区海洋生物多样性、渔业资源及其开发利用提供理论依据及数据支持。
1 材料与方法
1.1 研究区域及资料来源
现场调查数据来源于中国第四次北极科学考察(2010年7月1日—9月20日)。该航次调查船为中国“雪龙”号科学考察船,根据不同纬度在白令海共布设21个站位,分布在白令海中央海盆、白令海大陆坡、白令海大陆架以及白令海峡(58°00.00′N~64°20.14′N,176°12.24′E~167°20.52′W),各站位拖网水深分布范围为24~626 m。现场采样分别使用法式底拖网(宽2.5 m,高0.5 m,长9 m,囊网网目10 mm)、三角网(宽2.2 m,高0.65 m,长6.5 m,囊网网目20 mm)和阿氏网(宽1.6 m,高0.5 m,长3 m,囊网网目20 mm)进行底层拖网,使用IKMT网(长9 m,网口面积4 m2,囊网网目10 mm)进行中层拖网,拖网速度为2.4~3.9 kn;拖网时间视海况和底质情况有所变动,为8~45 min(见图1)。样品采样及分析严格按照《海洋调查规范》进行[25],鱼类鉴定及分类参考《Fishes of Alaska》和《Fishes of the World(4th)》[26-27]。
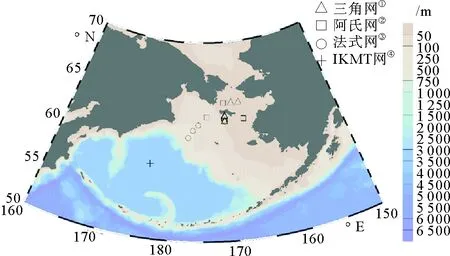
(①Triangular beam trawl;②Otter trawl;③French-type beam traul;④Isaacs-kidd midwater trawl.)
图1 中国第四次北极科学考察白令海站位图
Fig.1 Stations in Bering Sea during the 4th Chinese National Arctic Research Expedition in 2010
各海域鱼类物种资料来源于Fishbase[13]和Global Species[14]等数据库,按Nelson分类系统[27]系统整理了各海域鱼类物种组成,本研究中鱼类物种组成只选用鉴定到种的种类,学名主要参考《Fishes of the World(4th)》和WoRMS[28]。
1.2 鱼类相对丰富度指数
本研究采用物种相对丰富度指数在科级和目级阶元上对各海域的鱼类物种组成进行分析,对比各海域鱼类物种组成的不同。相对丰富度指数计算式如下:
R=Si×100/D。
(1)
式中:Si为该海域第i科(目)鱼类的物种数目;D为相应鱼类在该海域所记录的科(目)数。
1.3 鱼类相似性系数
为了将白令海与其他海域的鱼类共有种数进行有效对比,本研究采用Czechanowski系数[29]计算各海域之间鱼类种级阶元的种类相似性。Czechanowski系数计算式为:
SI=2C/(A+B)。
(2)
式中:A为甲海域记录的鱼种数目;B为乙海域记录的鱼种数目;C为2个海域所共有的鱼种数目;SI为Czechanowski系数。当两海域所含鱼种完全相同时,Czechanowski系数为1,而2个海域所含鱼种完全不同时,Czechanowski系数为0。
1.4 分类学多样性指数
平均分类差异指数和分类差异变异指数基于物种之间不同分类等级的权重及所在分类系统中的路径长度,根据物种的有无对群落物种的多样性进行评估[16],其表达式为:
平均分类差异指数(Average Taxonomic Distinctness, AvTD):
Δ+= (∑∑i < jωij)/[N(N-1)/2]。
(3)
分类差异变异指数(Variation in Taxonomic Distinctness, VarTD):
Λ+= ∑∑i < j(ωij-Δ+)2/[N(N-1)/2]。
(4)
其中:ωij为第i和j个种类在分类系统数中的路径长度,N为种类数。由于鱼类都属于脊索动物门,因此本研究中的分类阶元确定为纲、目、科、属、种,共5个分类阶元,不同分类阶元之间的加权路径长度ω的权重取值见表1[30]。分类多样性指数值由Primer5.0软件的TAXDTEST求得[31]。
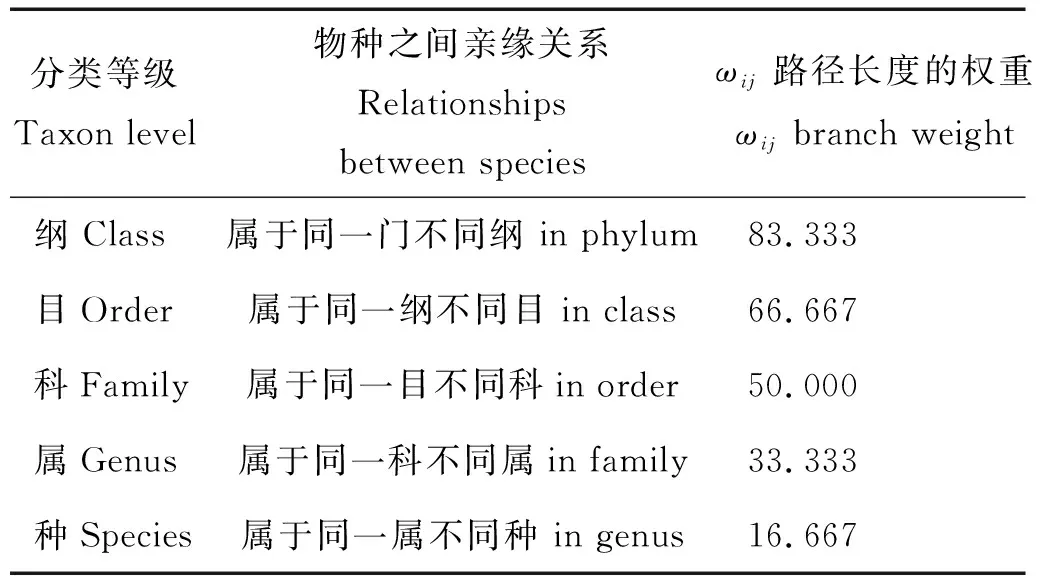
表1 分类等级多样性权重值
1.5 分类阶元包含指数
为了在各分类阶元(纲C、目O、科F、属G、种S)上表征各海域鱼类组成之间的多样性,本研究根据李娜娜等研究中所提出的分类阶元包含指数(TINCLi)[22],以分析鱼种各阶元分布的集中程度,其表达式为:
(5)
式中:Ni表示第i级分类阶元的数目;Ckj为第j个k级分类阶元的数目。
2 结果与分析
2.1 鱼类种类组成
2.1.1 本调查获取的鱼类种类及数量组成 中国第四次北极科学考察在白令海开展的21个站位调查中,共捕获鱼类31种(702尾),隶属于7目12科25属(软骨鱼类1目1科1属,硬骨鱼类6目11科24属),以鲉形目出现种类最多(13种,尾数占比21.5%),其次为鲈形目(10种,尾数占比29.2%)、鲽形目(4种,尾数占36.9%)、鳕形目、鳐形目、巨口鱼目及鲱形目各1种(尾数占比12.4%)(见表2)。

表2 2010年中国第四次北极科学考察获取的白令海渔获物种类组成
2.1.2 数据库记录的鱼类物种组成 由数据库记录的鱼类物种资料统计结果表明,白令海鱼类共359种,隶属于21目61科186属,其中头甲纲1目1科3属,软骨鱼纲3目4科6属,辐鳍鱼纲17目56科177属(见表3)。目级水平上,鲉形目物种数占绝对优势,共169种,隶属于11科64属;其次为鲈形目,共76种,隶属于13科37属;及鲽形目,共25种,隶属于3科18属。从科级水平上看,白令海以杜父鱼科种数最多,共60种;其次为狮子鱼科46种,鲽科23种及绵鳚科8种。

表3 资料记录的白令海鱼类物种组成
2.2 相对丰富度
由数据库记录的鱼类物种资料计算相对丰富度指数得出:目级水平上,白令海以鲉形目鱼类(804.8)占绝对优势,其次为鲈形目(361.9)、鲽形目(119.0)、胡瓜鱼目(81.0)及鳕形目(71.4)。科级水平上,鲉形目中的杜父鱼科(98.4)相对丰富度指数最高,其余较高的8科分别为狮子鱼科、绵鳚科、鲽科、线鳚科、八角鱼科、鲉科、圆鳍鱼科和鳐科,多为鲉形目下的种类(见表4)。
白令海、楚科奇海、阿拉斯加湾、鄂霍次克海、日本海、黄海及南海7个海域鱼类物种相对丰富度水平对比(见表5)结果显示,目级相对丰富度水平上,位于高纬度海域的楚科奇海、白令海、阿拉斯加湾及鄂霍次克海都是以鲉形目丰富度指数最高,其次为鲈形目;而中低纬度的日本海、黄海及低纬度的南海则是以鲈形目丰富度指数最高,鲉形目次之,且纬度越低鲈形目相对丰富度指数越大。科级相对丰富度水平上,楚科奇海、白令海、阿拉斯加湾、鄂霍次克海及日本海以杜父鱼科、绵鳚科、鲽科及线鳚科等冷水性种类丰富度指数占优,而黄海与南海的优势种类相似,排名前5位的科均为鲈形目鱼类,分别都是鰕虎鱼科、隆头鱼科、鮨科、雀鲷科和天竺鲷科,多数种类为热带及亚热带的岩礁性鱼类,具有典型的热带及亚热带性。此外,成对比较Kruskal-Wallis一致性检验结果表明,鲈形目鱼类科级相对丰富度水平上,白令海与楚科奇海、阿拉斯加湾、鄂霍次克海和南海差异不显著(P>0.05),而与日本海和黄海的丰富度差异显著(P<0.05);鲉形目鱼类科级相对丰富度水平上则与楚科奇海、阿拉斯加湾、鄂霍次克海和日本海差异不显著(P>0.05),而与黄海和南海的差异显著(P<0.05)(见表6)。
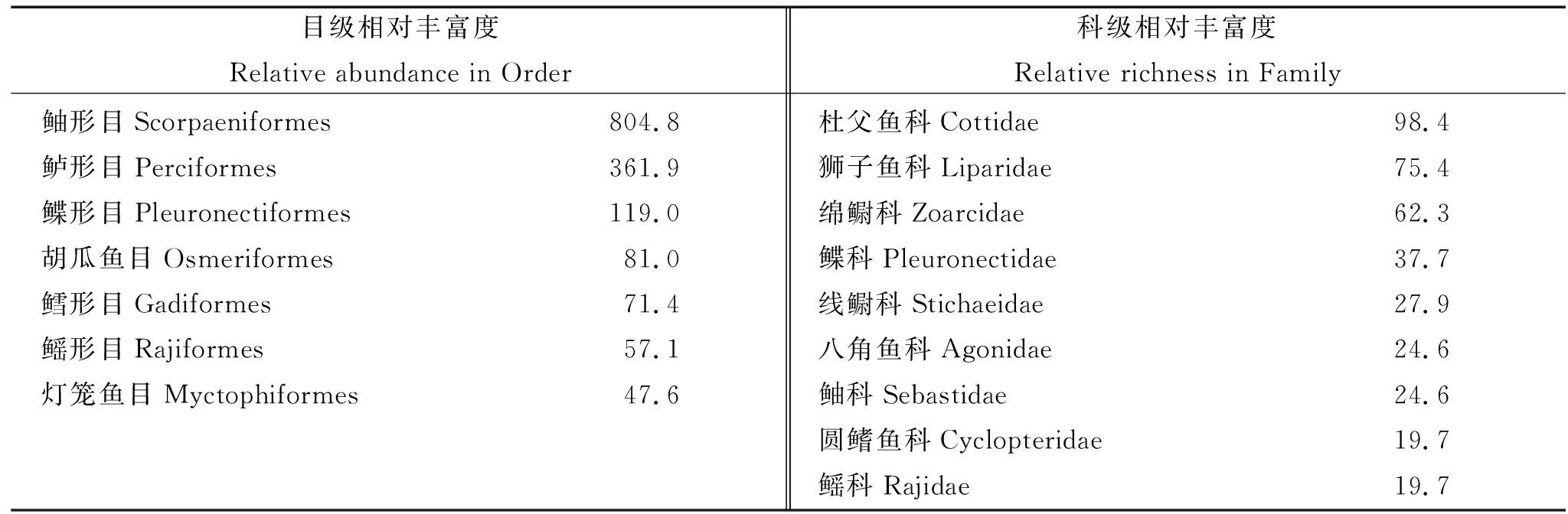
表4 白令海相对丰富度指数

表5 各海域相对丰富度靠前的目和科
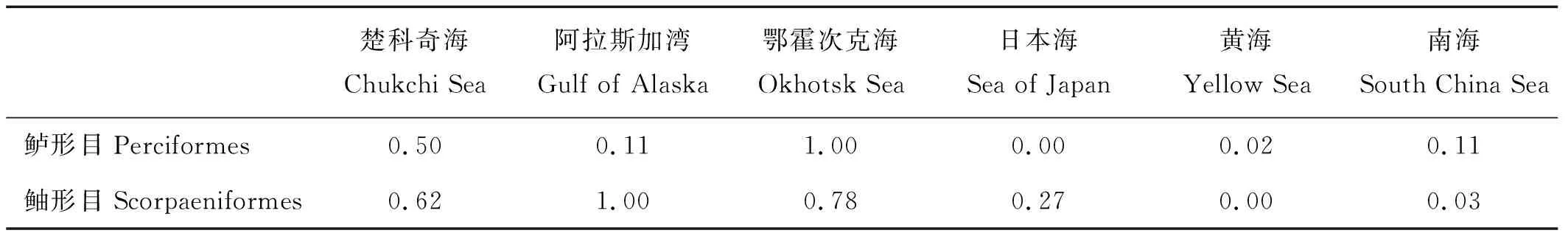
表6 白令海与其他海域鱼种相对丰富度差异Kruskal-Wallis检验P值
2.3 鱼类物种相似性系数
本研究白令海与其他中高纬度海域间共有种及相似性系数的对比显示(见表7),白令海与阿拉斯加湾的共有种数目(178种)及相似性系数(0.521)均为最高;其次为鄂霍次克海(共有种数为166种,相似性系数为0.437);与楚科奇海相似性系数较低(0.278),但共有种数(63种)占到楚科奇海总鱼种数目的67.0%。
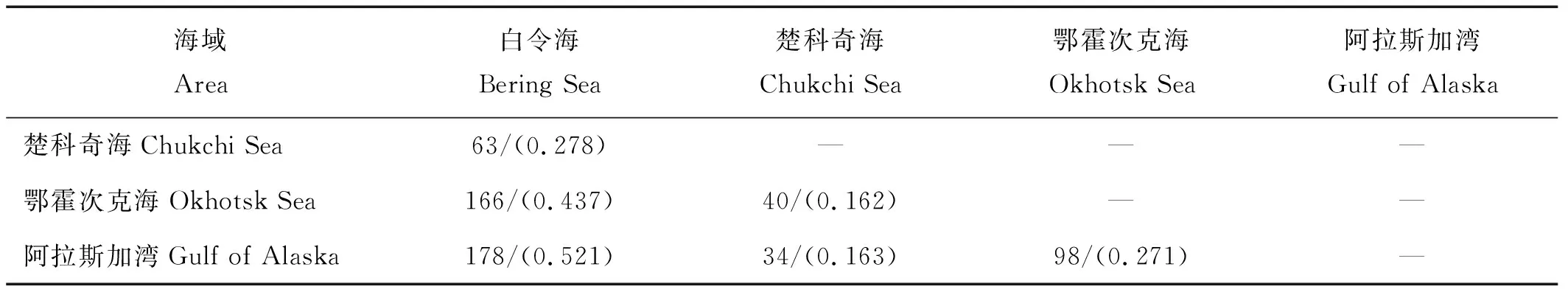
表7 各海域鱼种共有种数及相似性系数
2.4 分类学多样性
白令海鱼类物种组成分类阶元包含指数的分析结果显示,白令海鱼类目级阶元拥有(科、属、种)的数目为(2.90;8.86;16.48),科级阶元拥有(属、种)的数目为(3.05;5.67),属级阶元拥有(种)的数目为(1.80);分类多样性指数的分析结果显示,白令海平均分类差异指数△+为62.7,分类差异变异指数∧+为185.0,图2、3为白令海鱼类平均分类差异指数及分类差异变异指数的理论平均值(虚线所示)及95%的置信区间。

图2 白令海鱼类平均分类差异指数△+
将各海域鱼类分类多样性指数及分类阶元包含指数进行对比(见表8),结果表明,7个海域的平均分类差异指数△+相距不大,日本海△+稍高于其他6个海域,表明日本海海域鱼类物种间分类上差异稍大。分类差异变异指数∧+呈现出纬度差异,高纬度海域要高于低纬度海域,表明在高纬度海域鱼类物种组成亲缘关系分布更加不均匀。分类阶元包含指数S/G及S/F除了日本海为异常低值外,其余海域的数值都随着海域纬度的降低呈现增大的趋势,处于热带海域的南海S/O就显著高于楚科奇海的S/O。
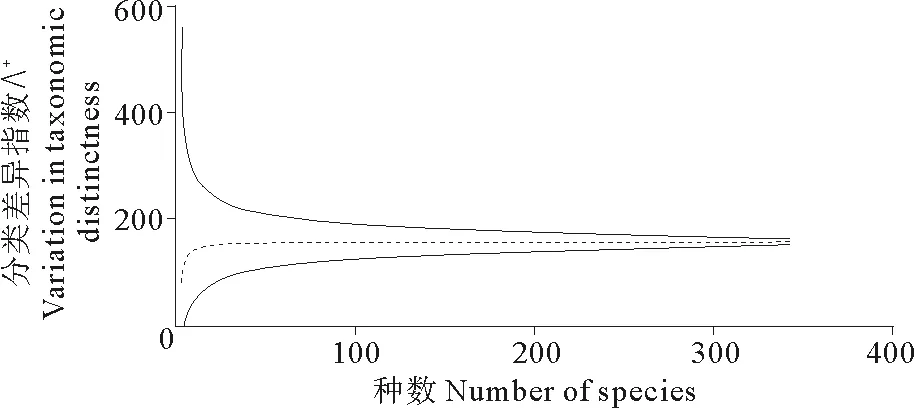
图3 白令海鱼类分类差异变异指数∧+Fig.3 Variation in taxonomic distinctness of fish in Bering Sea
3 讨论
3.1 鱼类物种组成特征
由数据库记录的鱼类物种统计资料来看,科级水平上白令海以杜父鱼科、狮子鱼科和鲽科种类数占优,其中前两科隶属于鲉形目,鲽科隶属于鲽形目。朱元鼎等对杜父鱼科和狮子鱼科等鲉形目鱼类的区系特征研究显示,除了细纹狮子鱼(Liparistanakae)、淞江鲈(Trachidermusfasciatus)等少数几种能分布至我国沿海以外,其他属种均分布于北太平洋高纬度冷水海域[32];成庆泰对鲽形目鱼类区系特征的研究表明,鲽科鱼类中的多数种类为中高纬度海域冷水性或冷温性种类[33]。因此,白令海鱼类组成应以冷水性和冷温性种类为主,对中国第四次北极科考渔获物的鱼种适温性分析结果与数据库资料分析结果一致,白令海捕获的31种鱼类均为冷水性或冷温性种类[6, 34]。

表8 各海域分类多样性指数及分类阶元包含指数对比
本研究中各海域鱼类相对丰富度对比结果显示,目级相对丰富度水平上白令海以鲉形目(804.8)为最高,随着海域纬度的降低,鲈形目鱼类逐渐增加,鲈形目相对丰富度水平上白令海(361.9)远小于黄海(2 665.1)和南海(4 358.3),表现出在暖温带及热带海域鲈形目鱼类种类数目的绝对性优势。从科级相对丰富度水平上看,白令海与楚科奇海、阿拉斯加湾及鄂霍次克海相近。在日本海,鰕虎鱼科、鲹科、鮨科等温带、亚热带的鱼类都已有出现,呈现出典型的暖温带与冷温带过渡特性。出现这种鱼类组成差异的原因与各海域的水文条件以及海流对物种的输送有关:源于黑潮分支和东海陆架水的黄海暖流和济州暖流,能延伸至35°N~36°N的黄海中部[35-36],导致位于暖温带的黄海海域,其科级相对丰富度排在前5的科与位于热带海域的南海相同;白令海峡以富营养的太平洋水团向北输送为主,对整个北极海域的水流交换和营养供应起至关重要的作用[48-49],因此白令海与楚科奇海鱼类共有种占楚科奇海鱼种数比例较高。而随着全球气候变化导致的白令海冷水团北移、海水表面温度上升等[8, 37],已经引起了部分鱼类向高纬度移动[6, 34],这可能造成白令海鲈形目鱼类种类数有所增加,从而改变白令海的鱼类物种组成结构。
3.2 分类多样性特点
本研究中除了白令海和阿拉斯加湾纬度覆盖范围有较大重复外,其余各海域鱼类物种数均随着纬度的降低而增加,但平均分类差异指数△+较为接近,即白令海与本研究其他各海域之间,平均分类路径长度并没有因为种类数多寡而发生较大变化。相反,分类差异变异指数∧+除鄂霍次克海高于白令海和阿拉斯加湾外,其余海域∧+随着纬度的增高而增高,即白令海等高纬度海域比低纬度海域鱼类亲缘关系更加多样化。从分类阶元包含指数上也能看出这一点,除了日本海S/G、S/F异常低外,其余分类阶元包含指数同样表现出了明显的纬度梯度变化规律,即随着纬度的升高,各分类阶元包含指数减少,表现在热带低纬度海域在同一目、同一科及同一属内的平均物种数目要远高于高纬度海域,近缘种较多,这与Shin等、Ellingsen等、陈国宝及李娜娜等的研究结果一致[22, 38-40]。全球海洋生物物种数目大致以印度洋—菲律宾海域为中心沿纬度变化而呈梯度递减分布,这一现象深层次的原因可能是,低纬度海域物种遗传物质变异速率较快,促进了物种的分化,使得低纬度海域新物种的形成速率要远大于高纬度海域,且随着离印太“珊瑚三角”的距离越远,物种分化并存在于地球上至今的年限越久远,物种越古老[41-42]。日本海S/G和S/F的异常低值,可能与该海域独特的环流特点有关,日本海作为半封闭的海盆,其具有独立的深层环流系统,仅通过4个浅而狭窄的海峡与外海相通,水交换相当有限[43-44]。
3.3 白令海鱼类区系划分
世界海洋鱼类多源于太平洋的两个源区,一是位于巴布亚新几内亚—印度尼西亚—菲律宾的海洋生物多样三角,其为暖水性鱼类的发源地;另一源区为北太平洋,是鲱科、鳕科、鲽科和鲉科等冷水及冷温性鱼类的发源地[51]。Briggs的研究也同样显示了诸如鲑科、绵鳚科、杜父鱼科、八角鱼科、狮子鱼科及线鳚科等鱼类起源于北太平洋冷水区[52],这与本研究中白令海此类鱼种科级相对丰富度水平较高的结果相吻合,而如鰕虎鱼科种类一般随着纬度的增加而减少,热带多于温带,寒带极少,极地及深海几乎无分布[53]。经过海域间的物种扩散,白令海的鱼类南可通过日本海到达中国黄、渤海海域,北可穿过白令海峡进入北冰洋以至北大西洋,如太平洋鲱(Clupeapallasii)、秋刀鱼(Cololabissaira)及大麻哈鱼(Oncorhynchusketa)等冷水性及冷温性鱼类在日本及朝鲜产量甚高;而狭鳕(Gaduschalcogrammus)、太平洋鳕(Gadusmacrocephalus)等在黄海也有分布[54]。中日两国冷水性及冷温性鱼类分布格局是同源扩布的结果[51],其源区即为北太平洋冷水性鱼类的发源地,但由于海域的阻隔及海流的影响,黄海海域冷水性及冷温性鱼类仅在中部黄海冷水团的保护下才少部分扩散并栖息至此。
海洋生物地理学的划分主要是根据物种的分布范围进行[45],而由此衍生出诸多海洋生物地理学的划分系统,如Briggs 区系省、Longhurst生物地球化学区系省及大海洋生态系(LMEs)等,其中,海洋区系省(Marine Provinces)是指主要受到海域地形、水文、地理化学等因素的影响,生物在进化上保留了一定的核心种类又兼具一些地方种类的海域[46]。据于此,Briggs将白令海分为3个区系:首先白令海北部是极地区系的一部分,极地区系通过白令海峡向南延伸至白令海的Olyutorsky角(Cape Olyutorsky)和努尼瓦克岛(Nunivak Island),南边界线是白令海1—2月份平均浮冰线;其次白令海西南部由Olyutorsky角沿勘察加半岛、千岛群岛岛链至日本本州岛和北海道岛之间的津轻海峡为千岛区系省(Kurile Province),它和东方区系省(Oriental Province)、鄂霍次克区系省(Okhotsk Province)归为西北太平洋冷温带区系;白令海其余部分为阿留申群岛区系省(Aleutian Province),属于东北太平洋冷温带区系[47]。Briggs的这种划分与本研究关于白令海与鄂霍次克海、阿拉斯加湾等具有较高种类相似性系数的研究结果相一致(见表4),其共有种相似性系数分别为0.437和0.521,这可能与白令海和阿拉斯加湾、鄂霍次克海有强大的海流进行水交换有关[50]。
4 结语
白令海是对气候变化最敏感的区域之一,人类活动同样对于白令海及北极海域的生态环境及渔业资源带来了较大影响,本研究利用Fish base及Global Species鱼种数据库资料,并结合第四次北极科学考察在白令海所得到的鱼类现场调查资料,首次利用分类学多样性方法研究白令海鱼类组成及分类多样性水平,从而跟踪白令海鱼类种类组成和区系变化,对研究气候变化和人类活动对生物的影响极为重要。本文由于实地调查资料有限,海区鱼种组成有待进一步补充,对于白令海及亚北极地区海域生物及资源变化的监测将有助于我们对此类海区资源的进一步了解。
致谢:感谢第四次北极科考期间众多科考队友以及雪龙船全体船员在鱼类采样过程中给予的协助。
猜你喜欢
热带作物学报(2022年9期)2022-10-17
云南农业科技(2021年6期)2021-12-30
科学养鱼(2020年8期)2020-12-20
智富时代(2019年8期)2019-09-23
智富时代(2019年8期)2019-09-23
小天使·一年级语数英综合(2019年8期)2019-08-27
农村百事通(2019年2期)2019-03-26
湖北农业科学(2017年19期)2017-11-09
摄影之友(影像视觉)(2017年1期)2017-07-18
青春岁月(2017年1期)2017-03-14
