
外来植物三叶鬼针草的入侵机制及其防治与利用
2019-02-19 02:23:24尚春琼朱珣之
草业科学 2019年1期
尚春琼,朱珣之
(1. 江苏科技大学生物技术学院,江苏 镇江 212018;2. 江苏省中国科学院植物研究所,江苏 南京 210014)
三叶鬼针草(Bidens pilosa),菊科(Compositae),鬼针草属(Bidens),一年生草本植物,原产于中美洲,目前亚洲和美洲的热带及亚热带地区均有分布,其瘦果冠毛芒状,具倒刺,可粘附于人畜、货物进入我国境内,1857年首次被报道,现广泛分布于我国华东、华中、华南、西南等地[1]。
三叶鬼针草入侵性和环境适应力强,其成体植株对光、温度、氮素有较强的表型可塑性,且产种迅速,结实量大,种子萌发率高,这些特性使得三叶鬼针草扩散到一个新生境后,在一到两代内就能产生一个大的种群,从而快速完成定殖和入侵。土壤微生物与三叶鬼针草的协同作用加快了三叶鬼针草的入侵,还可降低干旱胁迫对三叶鬼针草的伤害。三叶鬼针草入侵当地生态系统后,其强烈的入侵性除了因其适应性、繁殖能力极强而表现出争夺本地物种光照、水分、营养外,还常以化感作用直接或间接地危害其他物种[2]。
国内外现有关于三叶鬼针草的研究大多集中在三叶鬼针草的药用价值及重金属富集方面[3-5],对于三叶鬼针草的入侵机制和化学成分研究较少。尽管研究者[6]普遍认为三叶鬼针草的成功入侵与其产生的化感物质存在必然联系,但现有的研究并未真正揭示其化感作用机制。本研究从三叶鬼针草的生物学特性、入侵机制、入侵的生态学效应、防除及利用这5个方面对三叶鬼针草的研究做一总结和展望,希望对以后的研究有所帮助。
1 三叶鬼针草的生物学特性
1.1 形态特征
三叶鬼针草瘦果长条形,略扁稍弯,黑褐色;果体长7~10 mm,宽约0.8 mm;表面具有疏或密的暗黄色瘤和4条强劲纵棱,棱间有时具一细棱,棱上部具有向上的毛状短刺。顶端果针2~4枚,3枚者则中枚短,由腹棱伸出,针体具2列倒刺,基端果脐黄色,向背面倾斜,椭圆形凹陷[2]。
1.2 繁殖特性
三叶鬼针草为六倍体(2n = 72),具有较强的繁殖力和散布力。三叶鬼针草的交配机制灵活,不仅能异交传粉,还可自交结实,单个花序内可自交亲和。其花粉胚珠比 (P/O)为 1 754.12 ± 29.87,从开花到果实成熟只需18 d,种子成熟后自然脱落;若以单个植株500个花序计算,每株可产生约18 115粒种子[7]。三叶鬼针草种子一年四季均可成熟,其中春、夏季成熟占14%,秋季成熟占50%,冬季成熟占36%;发芽的主要季节则是春、夏季(约占66%)[1]。三叶鬼针草种子具有多种休眠机制,比如对红光和后熟的要求。Whitaker等[8]发现,三叶鬼针草中的活性氧类可以缓解其种子的休眠,·OH可代替后熟,O2·-可以部分取代红光。
1.3 生理生化特征
三叶鬼针草对光变化具有较强的适应性。无论是在强光还是光照不足的条件下,三叶鬼针草均表现出较高的资源利用率。光照充足时,三叶鬼针草比本地种金盏银盘(Bidens biternata)有更高的叶面积、生物量和株高[9];比本地种小蓟(Cirsium setosum)有更高的气体交换系数、叶绿素含量以及叶片氮、磷浓度,能够更有效地捕获和利用环境资源[10]。当光强受限时,三叶鬼针草通过增加比叶面积,降低叶片构建成本,提高光能捕获率,在竞争中获得比本地种更多的光资源[9]。有研究发现,光强受限时,三叶鬼针草繁殖期的叶片数比营养期有所增加,通过增加叶片数来获取更多的光源[11]。对于不同光强,三叶鬼针草还会在资源分配和能量积累方面做出适应其生长的可塑性调节,中度遮阴可促进其支持结构的生长和营养期的物质积累,而充足的光源能使繁殖期积累更多物质,为繁殖后代做准备[11]。
三叶鬼针草对温度的适应范围广,对土壤氮素响应的表型可塑性较大。15~30 ℃条件下,三叶鬼针草种子的萌发率均在80%以上[12];高温40 ℃的萌发率仍有46.5%,是土著种鬼针草(B.bipinnata)的9倍,而在低温10 ℃时,三叶鬼针草的萌发率也达到68%[13]。此外,经4 ℃和-10 ℃贮藏6个月的三叶鬼针草种子,其萌发率与贮藏前的种子相比无显著差异[14]。王瑞龙等[15]发现,全球变暖引起的温度升高可使三叶鬼针草幼苗增加对茎和叶的生物量投资,显著提高三叶鬼针草的株高和叶面积,同时增强三叶鬼针草对受体植物的化感作用。三叶鬼针草对土壤氮素响应的表型可塑性较大,表现在形态、生物量分配等方面,比如分枝数、总叶面积、叶片数的表型可塑性指数较高。当土壤中氮肥贫瘠时,三叶鬼针草减少叶片和分枝数的投入,加大对地下根生物量的投入,从而依靠根的吸收来竞争地下营养资源。而土壤中氮素充足时,三叶鬼针草通过增加叶片数和总叶面积来提高地上部的物质投入,增强光合作用,加大竞争和利用光能的能力[16]。
除了对光、温度和氮素有较强的可塑性外,三叶鬼针草对干旱胁迫也有较高的耐受性,种子萌发对pH的适应范围也较广,只有pH 2.0的强酸性溶液才会造成其萌发率显著下降[13]。还有研究发现,三叶鬼针草可以抵抗其他植物的化感作用,当其生长于紫茎泽兰(Eupatorium adenophorum)重度入侵地土壤时,三叶鬼针草的生长不受影响[17],且紫茎泽兰枯落物的水浸液还能提高三叶鬼针草的长势,促进三叶鬼针草体内物质的积累[18]。
2 三叶鬼针草的入侵机制
2.1 与土壤微生物协同作用
丛 枝 菌 根 真 菌 (arbuscular mycorrhizal fungi,AM真菌)是自然界中最广泛的植物共生真菌,能提高宿主对非生物逆境的抗性,尤其是抗旱性[19-21]。它可以影响宿主植物三叶鬼针草根系的分枝状况,促进其生长,使宿主能够吸收更大空间内的水分和营养物质。王宁和秦艳[22]用WinRHIZO图像分析系统和 Fractal analysis system for Windows软件分析了AM真菌对三叶鬼针草根系形态的影响,结果发现,接种AM真菌后的三叶鬼针草,其生物量、根系总长度、根系体积、根系表面积、根系分枝强度和细根 (d<0.5 mm)所占比例均显著增加。此外,AM真菌能够改善干旱胁迫条件下三叶鬼针草的气孔导度和羧化效率[23],且AM真菌的外延菌丝还能吸收到非菌根植株根系摄取不到的水分[24],从而进一步减弱水分亏缺对三叶鬼针草的伤害,但土壤水分匮乏严重或土壤干旱时间较长时,AM真菌对三叶鬼针草的这种保护作用会受到限制。三叶鬼针草在不同入侵程度土壤中还通过改变土壤细菌群落的组成和种类,打破当地植物与土壤微生物之间长期共生关系的平衡,降低当地植物的竞争力,并营造有利于自身生长的微环境,以便进一步扩张[25]。
2.2 化感机制
三叶鬼针草的化感作用可以有雨雾淋溶、叶片挥发、根系分泌等方式(表1)。从化感物质的淋溶途径出发,三叶鬼针草水浸提液可显著抑制莴苣(Lactuca sativa)、 香 椿 (Toona sinensis)、 龙 须藤(Bauhinia chmpionii)和苏木(Caesalpinia sappan)4种岩溶木本植物的种子萌发、幼苗生长和光合作用[6,26],降低蒲公英(Taraxacum mongolicum)种子的发芽势、发芽率,提高体内丙二醛含量及相对电导率,造成蒲公英膜质过氧化程度加大,膜渗漏加剧[27]。在禾本科植物中,高浓度的三叶鬼针草水浸提液会使小麦(Triticum estivum)幼苗的根变黑、腐烂[26]。经显微分析发现,受三叶鬼针草水浸提液处理后的小麦根尖,其细胞内染色体出现微核、染色体断片、染色体桥等多种畸变[28]。从化感物质的气体挥发途径来看,三叶鬼针草对受体植物的影响会因挥发物浓度不同而有所差异,其化感作用表现为促进和抑制两种形式。5 g三叶鬼针草叶片所释放的挥发物,可诱导旱稻(Oryza sativa)初生根髓腔形成,促进维管柱的发育,但20 g三叶鬼针草叶片的挥发物则会引起旱稻幼苗初生根结构变异,维管柱消失,髓腔变大[29]。三叶鬼针草除了通过地上部分释放化感物质影响邻近植物生长外,也会利用地下根系的分泌物对周围的种子植物产生化感作用。研究发现,三叶鬼针草根系分泌物可抑制蕨类植物井栏边草(Pteris multifida)配子体生长和发育,降低其光合作用能力[30],高体积分数的根系分泌物还可对配子假根造成损伤,引起细胞破裂和死亡,配子致死率高达88.3%[31]。

表1 三叶鬼针草的化感作用Table 1 Allelopathic effects of Bidens pilosa
3 入侵的生态效应
三叶鬼针草对入侵生境的土壤生态系统结构和功能存在影响,且三叶鬼针草的不同发育阶段产生的影响不同。何兵等[32]发现三叶鬼针草果期对土壤酶活性的影响大于苗期,果期根际土壤中的脲酶、蔗糖酶、酸性磷酸酶等活性显著升高,且土壤理化性质发生改变,有机质含量显著升高。分泌土壤酶是土壤微生物发挥其生态功能的一种途径。三叶鬼针草也可通过改变土壤微生物群落结构间接影响土壤酶活性。三叶鬼针草入侵后,其根际土壤中的细菌和真菌数量显著增加,这种改变提高了土壤中氮、磷含量,加速土壤养分循环,从而增强了三叶鬼针草吸收氮、磷的能力[25,32]。但也有研究发现,三叶鬼针草变种(B . pilosa var. radiate)的挥发油对6种细菌[黄色微球菌(Micrococcus flavus)、枯草芽孢杆菌(Bacillus subtilis)、蜡样芽胞杆菌(B.cereus)、短小芽孢杆菌(B.pumilus)、大肠杆菌(Escherichia coli)、假单胞菌(Pseudomonus ovalis)]和3种真菌[腐皮镰刀菌(Fusarium solani)、尖孢镰刀菌(F.oxysporum)、罗尔伏革菌(Corticium rolfsii)]显示出抗细菌和抗真菌活性[33]。芽孢杆菌常被用来作为抑制植物病原菌的生物农药使用,而芽孢杆菌的抑菌作用可能对当地植物的生长起到促进作用[25],三叶鬼针草挥发物对芽孢杆菌起到抑菌效果,所以间接地降低了当地植物的竞争力,帮助三叶鬼针草成功入侵。
4 三叶鬼针草的防治
4.1 化学防治
郭成林等[34]研究了8种除草剂对木薯地三叶鬼针草的防除效果,其结果发现,500 g·L-1特丁噻草隆对三叶鬼针草的防除效果最佳,其次为60%的恶草·丁草胺和57%的氧氟·乙草胺。另外50%丁草胺、90%乙草胺、960 g·L-1精异丙甲草胺、120 g·L-1恶草酮、24%乙氧氟草醚5种除草剂对三叶鬼针草的防除也具有一定效果。宣红燕[35]提出防治路边或高大乔木林里的三叶鬼针草时,使用百草枯100倍液喷施效果较好,而防治农作物中的三叶鬼针草,可与100~200倍液的增效剂乙草胺联用。但由于百草枯使用不当对人体造成的伤害极大,且无特效解药,所以自2016年7月1日起,百草枯水剂在我国已被禁用。甲磺隆为现有磺酰脲类除草剂中活性最高的品种,已被应用于入侵植物紫茎泽兰和加拿大一枝黄花(Solidago canadensis)[36]的防除,在三叶鬼针草三叶期时连续两年4次施用浓度为8 mg·L-1的甲磺隆,三叶鬼针草的防除效率为100%[37]。与甲磺隆一样, 草甘膦也是一种灭生性化学除草剂,它主要是通过茎叶吸收后传导到植物组织,影响蛋白质合成,最终导致植物死亡[38]。经草甘膦处理后的三叶鬼针草种子,其发芽率显著降低,幼苗质膜完整性遭到破坏,但为了抵御草甘膦带来的氧化损伤,三叶鬼针草会提高保护酶活性[39]。虽然导致目标植物死亡使甲磺隆和草甘膦在防除效率上具有较高的优势,但这种特性也会对环境造成破坏,所以不建议使用灭生性除草剂对三叶鬼针草进行防除。
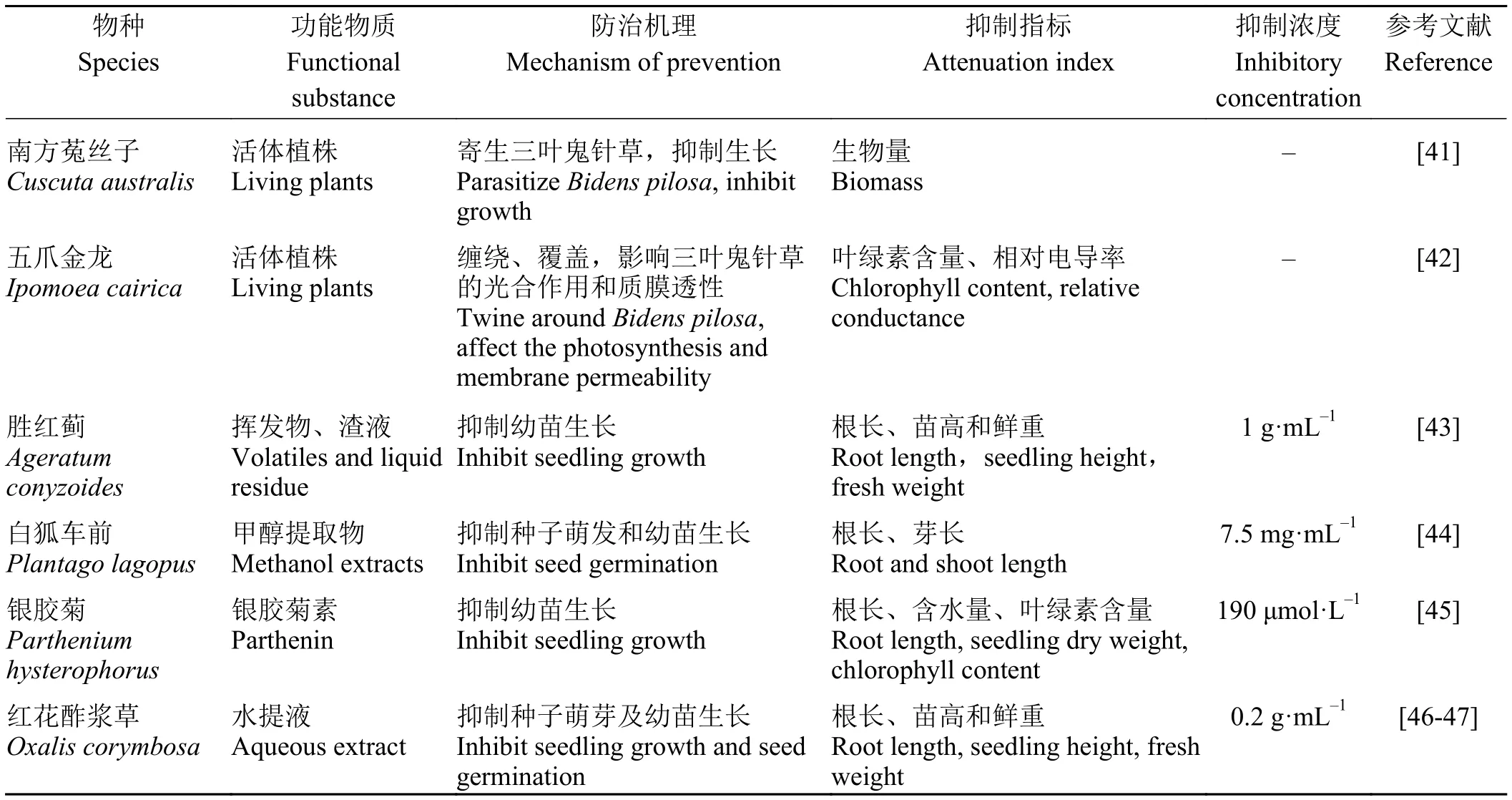
表2 三叶鬼针草的生物防治Table 2 Biological control of Bidens pilosa
4.2 生物防治
生物除草剂是指利用自然界中的生物(包括微生物、植物和动物)或其组织、代谢物工业化生产的用于除草的生物制剂,主要分为两类。一类是直接用完整生物体或部分活体组织开发的生物制剂;另一类是利用生物的次生代谢产物开发的生物源除草剂或生物化学除草剂[40]。经文献整理发现,用于三叶鬼针草的真菌或细菌生物除草剂还未有报道,但发现两种可抑制三叶鬼针草生长的植株,以及多种具有植物源生物除草剂开发潜能的入侵植物(表2)。
针对三叶鬼针草入侵后形成的植物群落,可以利用入侵地的当地寄生植物对三叶鬼针草进行寄生,影响三叶鬼针草的生长和繁殖,促进当地植物的生长,进而改变三叶鬼针草的群落结构和多样性,达到生物防治及生态恢复的目的[48]。张静等[41]发现南方菟丝子(Cuscuta australis)寄生三叶鬼针草后,其缠绕圈数、吸器数等都随寄生时间的延长而增加,34 d后可显著抑制三叶鬼针草的生长,且用两段15 cm的南方菟丝子茎段寄生三叶鬼针草幼苗最有效。此外,五爪金龙(Ipomoea cairica)是缠绕藤本,茎柔软,盘绕能力极强,三叶鬼针草作为它的伴生种,在它的盘绕、覆盖下,三叶鬼针草的光合作用能力低下、质膜受损[42]。但五爪金龙也属于入侵杂草,若将其用于三叶鬼针草的防治,可能会对修复地造成二次入侵。
植物源生物除草剂大部分来源于植物的化感物质,主要是一些有机酸、酚类、生物碱、类固醇、单宁、类萜、甾类化合物和醌类等[40]。研究发现,胜红蓟(Ageratum conyzoides)地上部挥发油及渣液对三叶鬼针草幼苗生长具有显著抑制作用,特别是在渣液浓度为1 g·mL-1时,三叶鬼针草种子的萌发受到完全抑制[43]。用 7.5 mg·mL-1的白狐车前 (Plantago lagopus)和 10 mg·mL-1的 大 车 前 (P.major)甲醇提取物处理三叶鬼针草,也能使种子萌发完全抑制;10 mg·mL-1的两种水提物对三叶鬼针草的萌发的抑制率也达到了60%以上[44]。除此之外,银胶菊(Parthenium hysterophorus)中的银胶菊素,红花酢浆草(Oxlis corymbosa)水提液对三叶鬼针草种子的萌发及幼苗生长同样起着抑制作用,试验证明,银胶菊素对三叶鬼针草的半数致死浓度 LC50约为 700 μmol·L-1[45],而浓度为 0.20 g·mL-1的红花酢浆草叶水提液对三叶鬼针草的化感抑制作用较强[46-47]。
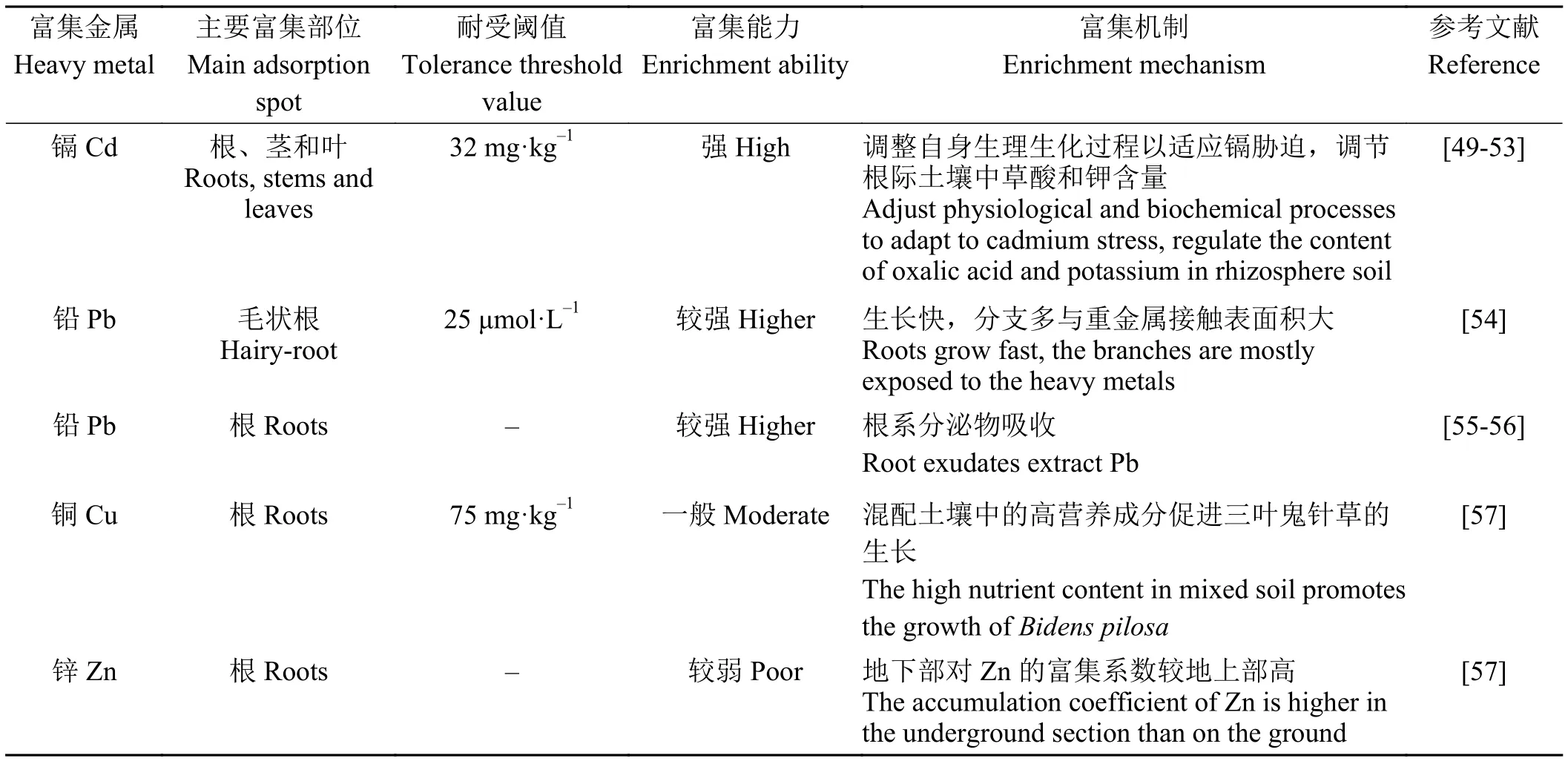
表3 三叶鬼针草的重金属富集作用Table 3 Enrichment properties to heavy metals
5 三叶鬼针草的应用
5.1 植物修复作用
超富集植物(hyperaccumulator)是指能够转移、超量积累一种或同时积累几种重金属元素的植物,超富集植物具备4个显著特征,一是富集重金属具有临界含量,镉为100 mg·kg-1;二是地上部分重金属含量必须大于其根部积累的重金属含量;三是植物对重金属必须具有较强的耐性,即重金属不会引起植物地上部生物量下降;四是地上部富集系数>l[4-5]。三叶鬼针草具备镉超富集植物的基本特征,是镉超富集植物(表3)。在三叶鬼针草的开花期和成熟期土壤中施以低浓度(≤16 mg·kg-1)镉时,三叶鬼针草不仅满足地上部镉含量大于100 mg·kg-1的条件,而且干重还有所增加[49],甚至在土壤镉浓度高达100 mg·kg-1时,三叶鬼针草地上部镉含量达到119.1 mg·kg-1[4]。那么,三叶鬼针草是如何耐受镉胁迫,并对镉进行吸收、积累的?研究表明,三叶鬼针草一是通过调整气孔导度、蒸腾速率,加强叶绿素合成,以保持光合作用的稳定进行;二是通过调节叶部脂肪酸组成来抵御镉胁迫引起的质膜氧化;三是改变渗透调节物质与抗氧化酶含量以维持渗透平衡和氧化损伤修复[50];四是通过调节根际土壤中草酸[51]和有效钾[52]含量来调节镉的迁移性和其自身的生物活性,从而调控根系对重金属的吸收与转运[50]。但需要注意的是,三叶鬼针草对镉的耐性存在阈值,当土壤镉浓度大于32 mg·kg-1时,虽然三叶鬼针草也满足富集系数大于1的条件,但其自身的生理生化过程会受到一定的影响,因此,修复土壤中镉浓度应尽量不超过 32 mg·kg-1[53]。
三叶鬼针草除了是镉超富集植物,还是砷排异植物(As-excluder)。当砷、镉复合污染时,三叶鬼针草表现出较强的耐受性,其地上部干重没有明显受到抑制,根、茎、叶中砷含量很低,富集系数和转移系数小,具备砷排异植物的基本特征[58]。但镉-砷复合污染土壤中砷的存在会抑制三叶鬼针草对镉的吸收,最强可使叶中镉积累量降低66%[59],所以三叶鬼针草最好用于非镉-砷复合污染土壤的修复。
三叶鬼针草的自然植株及发根农杆菌(Agrobacterium rhizogenes) C58C1 诱导的毛状根对铅污染土壤及水体有着较强的修复潜能[54]。三叶鬼针草的内源一氧化氮可以减少根中的活性氧,提高幼苗根系活力,减少铅胁迫造成的细胞膜氧化损伤[60]。处于重度铅污染土壤中的三叶鬼针草,其根系分泌物能够结合铅,完成铅的吸收[55]。复合污染土壤中,三叶鬼针草对铅的吸收和转运受其他重金属浓度、土壤参数和土壤细菌多样性的影响[56],其中含硫肥料的添加会降低三叶鬼针草对铅的吸收[61]。三叶鬼针草地下部对铜的富集系数大于1,在铜浓度低于75 mg·kg-1的污染土壤中能富集较多的铜,但地下部对锌的富集系数较地上部高,不满足富集植物的基本条件,所以可以结合拔式收割植物,从根部带走重金属离子,修复效果可能比其他生物量小的修复效果要好[57]。因此,三叶鬼针草可用作铜、锌污染土壤的植被重建材料,且土壤修复过程中可以通过添加有机酸改善地上部对锌的富集[51]。
5.2 饲料开发
三叶鬼针草营养价值和饲料转化率高,可用于开发草食动物饲料。邹知明等[62]采集广西野生的三叶鬼针草,晒制并加工、粉碎、制备成干草粉,添加制成配合饲料饲养家兔,试验6周后发现,三叶鬼针草组的家兔增重了 940 g。邓玲姣等[1,63]提出,若三叶鬼针草作饲料用,则适宜的刈割高度为61~80 cm,一年刈割3~4次,三叶鬼针草的总生物产量为 7.28 kg·m-2,干物质产量为 1.32 kg·m-2,粗蛋白质产量可达205.9 g·m-2;若用于商业生产,可通过脱叶、采花和疏果的方式延缓三叶鬼针草衰老,保持增长,增大可收获量[64]。
5.3 药用价值
三叶鬼针草属于我国民间常用草药,据唐代《本草拾遗》记载,三叶鬼针草具有清热解毒、散瘀消肿的功效[65]。现代仪器发现,三叶鬼针草的化学成分主要有黄酮类、多酚类、烯炔类、甾醇类萜类、有机酸及酯、香豆素类等[66-69]。药理学研究证实,三叶鬼针草具有降血糖、改善心力衰竭、抗炎、抗肿瘤、抗氧化、保护肝脏等作用(表 4)。
Chien等[70]发现,三叶鬼针草甲醇提取物可以显著降低Ⅱ型糖尿病小鼠体内糖化血红蛋白A1C(HbA1C)的含量,连续28 d口服三叶鬼针草水提物,也能使患病小鼠HbA1C水平显著降低,水提物能够刺激胰岛素分泌,保护胰岛结构,降低血糖含量[71]。Chen等[78]发现,三叶鬼针草中的多酚物质对肾素和血管紧张素转换酶(ACE)具有双重抑制作用,达到扩张血管、改善心力衰竭的效果。
三叶鬼针草对肠胃及肝脏具有保护作用。三叶鬼针草与类姜黄素的联合制剂可用于治疗5-氟尿嘧啶(5-FU)引起的小鼠肠粘膜炎,其作用是通过增加肠绒毛长度和隐窝深度来减少肠粘膜小鼠的病理学变化[72],调节髓过氧化物酶活性和抗炎性因子IL-10水平来缓解炎症[79];口服MMBP(只由绿肥栽培的三叶鬼针草加工成的粉末)能预防乙醇、吲哚美辛、冷刺激等引起的小鼠胃粘膜炎[80]。三叶鬼针草乙醇提取物可以降低幽门结扎大鼠的胃液、胃酸及胃蛋白酶的分泌量,抑制酒精诱发的胃出血[73]。三叶鬼针草中的总黄酮可以降低小鼠肝损伤程度,减轻大鼠肝纤维化病症。Yuan等[74]用三叶鬼针草总黄酮每天灌胃急性肝损伤大鼠,评估10 d后发现,TFB能有效降低大鼠血清谷草转氨酶(GOT)和血清谷丙转氨酶(GPT)活性,恢复肝脏超氧化物歧化酶(SOD)和谷胱甘肽过氧化物酶(GSHPx)活性。
在免疫系统调节方面,三叶鬼针草的甲醇提取物及分离出的聚乙炔成分(PA-1)可以阻断人和鼠淋巴细胞增殖,消减由酵母多糖免疫诱导引起的Pop淋巴结肿块[75]。乙醇提取物能够阻止人红细胞中谷胱甘肽、三磷酸腺苷的消耗以及SOD活性的下降,从而保护正常人红细胞免受氧化损伤[76]。三叶鬼针草的乙醇提取物和超临界萃取物还具有抗肿瘤活性,通过减小腹水和肿瘤细胞体积,延长荷瘤小鼠的存活时间[77]。100 μg·mL-1的三叶鬼针草水提物可显著抑制单纯疱疹病毒的复制[81]。
6 展望
三叶鬼针草作为外来入侵植物,依靠其强大的繁殖能力,在短期内占领环境空间,出现大范围的植被更替,严重破坏当地生态系统。但种子数量多、萌发快这一特性又使得三叶鬼针草作为饲料开发具有较好的利用价值。三叶鬼针草对环境因子变化具有强大的可塑性,比当地植物具有更强的资源竞争力,与土壤微生物的协同作用也帮助其入侵,三叶鬼针草的这些特性预警人们应对其预防和管理工作加以重视。
近年来,国外关于三叶鬼针草的研究多集中在其药用价值方面,而国内学者的研究则为三叶鬼针草作为镉富集植物的开发提供数据支持。然而,有关三叶鬼针草生殖、生长等基本生命特征的分子机理研究尚少,关于化学成分及化感作用分子机制的研究也鲜见。研究焦点都放在药用价值上,而忽视了其入侵性,这极易导致人们对三叶鬼针草的危害性认识不足,可能造成三叶鬼针草更大面积的蔓延。所以,三叶鬼针草今后的研究应平衡发展,加强对三叶鬼针草自身特性及入侵机制的研究。目前三叶鬼针草的防除主要通过人工铲除和化学防治,但这两种措施都只能在局部范围内起短期作用,不能较好地除去埋在土壤里的种子。当三叶鬼针草防除难度较大时,适当开展三叶鬼针草饲料及药用价值的开发研究是一种有效防除途径。利用当地物种的竞争作用或其他植物的化感作用抑制三叶鬼针草生长,具有防除三叶鬼针草的潜能,但此措施目前尚未成熟,有待深入研究。
猜你喜欢
儿童时代·幸福宝宝(2019年9期)2019-10-28 18:04:52
幼儿园(2018年15期)2018-10-15 19:40:36
现代园艺(2017年23期)2018-01-18 06:57:44
莫愁·家教与成才(2017年7期)2017-07-11 21:31:47
红领巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
新疆农垦科技(2016年10期)2016-06-15 20:29:33
杂草学报(2015年2期)2016-01-04 14:57:58
少儿科学周刊·儿童版(2015年10期)2015-11-07 03:42:03
少儿科学周刊·儿童版(2015年1期)2015-07-07 04:12:52
