
环渤海红条毛肤石鳖种群遗传多样性研究
2019-02-15 05:09王儒晓李媛媛刘传林杜文晓侯建海姚文君张俊凯张运发王宇曲江勇
四川动物 2019年1期
王儒晓, 李媛媛, 刘传林, 杜文晓, 侯建海, 姚文君, 张俊凯, 张运发, 王宇, 曲江勇
(烟台大学生命科学学院,山东烟台264005)
多板纲Polyplacophora是软体动物门Mollusca中较原始的类群,石鳖是多板纲的代表种类,从白垩纪至今,其化石形态变化不大,在研究软体动物的起源与进化中有重要的地位和意义(赵青松等,2007)。我国沿海分布10多种石鳖,以红条毛肤石鳖Acanthochitonrubrolineatus较为常见。红条毛肤石鳖分布于我国黄、渤海到广东沿岸,生活于潮间带和低潮区岩石岸,以附着性的藻类、底栖硅藻等为主要食物,属狭温、狭盐性动物(张均龙等,2015)。目前,红条毛肤石鳖的研究主要集中在形态、药用及磁性纳米材料(钱霞等,2015;王玲等,2018),其种群遗传结构与遗传多样性尚不明确。因此,开展红条毛肤石鳖不同地理种群的分子遗传研究,了解其遗传结构,对我国潮间带多板纲生物遗传多样性等研究具有重要意义。
线粒体细胞色素c氧化酶亚基Ⅰ(cytochrome c oxidase subunit Ⅰ,COⅠ)作为最常用的分子标记,在海洋生物的遗传多样性及种群遗传结构研究中发挥着重要作用(Nietal.,2012;Lietal.,2015;Corderoetal.,2017;王玲等,2018)。本研究利用线粒体COⅠ基因探讨环渤海红条毛肤石鳖9个地理种群的遗传结构,以期阐明环渤海红条毛肤石鳖的遗传多样性现状及其受环境因子的影响状况,为合理利用红条毛肤石鳖种质资源提供参考。
1 材料与方法
1.1 样品采集
采样点覆盖红条毛肤石鳖在渤海区域的全部分布区,基本涵盖全部岩基海岸潮间带,主要为低潮带。采样点分别是:莱州、营口、大连、葫芦岛、长岛、砣矶岛、烟台、秦皇岛、蓬莱(图1),共采集了环渤海海域9个样地126只红条毛肤石鳖个体,采集后保存于无水乙醇中备用。
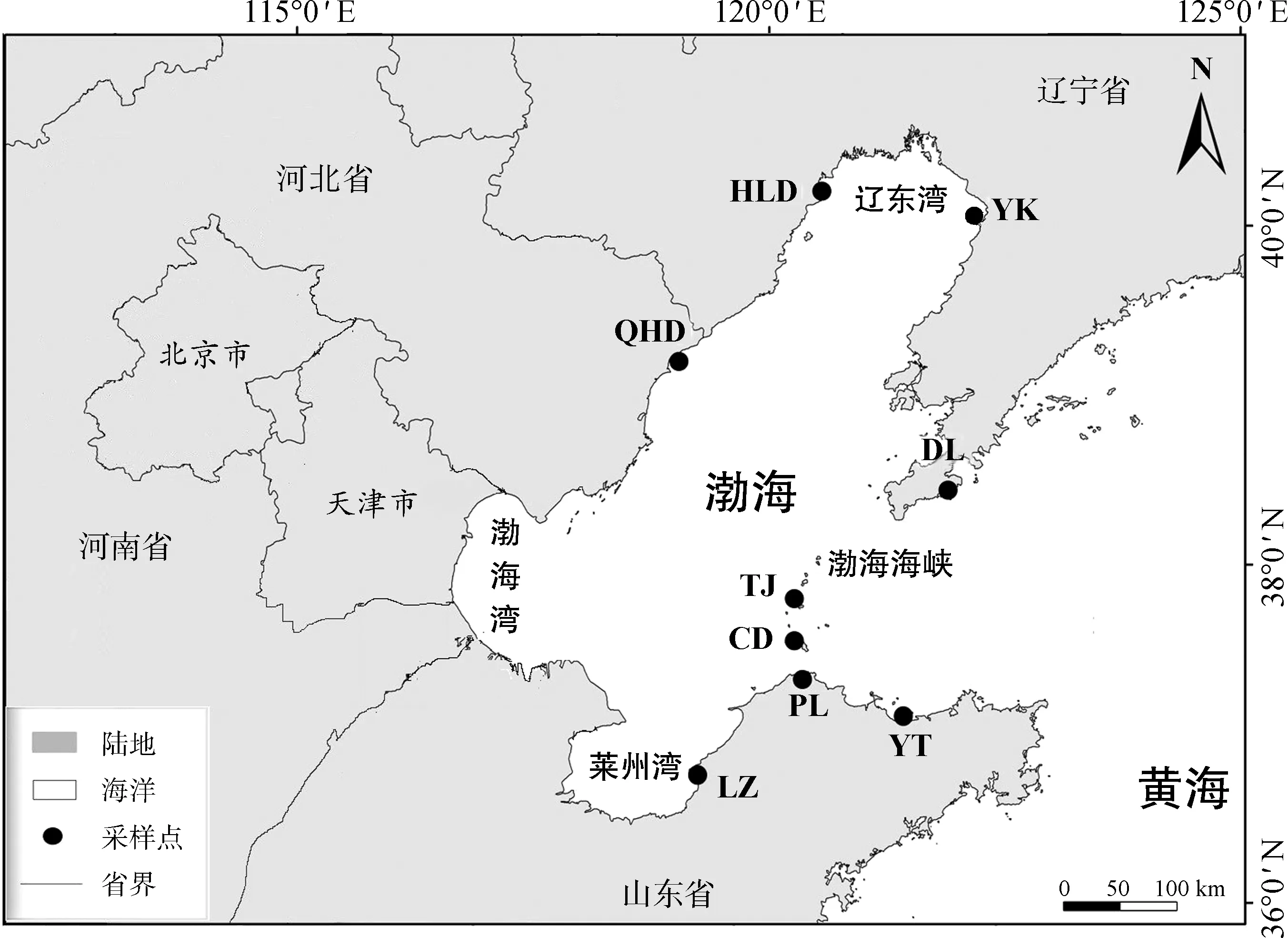
图1 红条毛肤石鳖地理种群采样点Fig. 1 Sampling sites of Acanthochiton rubrolineatus
DL. 大连Dalian (121°41′E, 38°54′N), YK.营口Yingkou (122°50′E, 40°37′N), HLD. 葫芦岛Huludao (120°57′E, 40°43′N), QHD. 秦皇岛Qinhuangdao (119°39′E, 39°56′N), TJ. 砣矶岛Tuojidao (120°46′E, 38°10′N), CD. 长岛Changdao (120°46′E, 38°10′N), LZ. 莱州Laizhou (119°56′E, 37°15′N), YT. 烟台Yantai (121°30′E, 37°27′N), PL. 蓬莱Penglai (120°49′E, 37°47′N); 下同the same below
1.2 环境因子的选取
选取经度、纬度、年平均温度、年平均温度变异系数、年平均降水量、年平均降水量变异系数、日照时数、日照时数变异系数、日照百分率变异系数、年平均水汽压变异系数、年平均气压、年平均气压变异系数共12个环境因子(图1、表1)。数据来源于国家气象数据共享服务网(http://data.cma.cn/),从该网站获取8个采样点近30年的气象数据并计算气温、降水量变异系数(砣矶岛数据未收录)。
1.3 DNA提取、扩增及测序
分别取红条毛肤石鳖肌肉约100 mg,采用DNeasy®组织&血液试剂盒(GTIN:0405322800-6077,Lot:151046088;GIAGEN,德国)提取总DNA。

表1 8个地理种群近30年环境因子(砣矶岛无数据记录)Table 1 Environmental factors of 8 populations in 30 years (no record in Tuoji Island)
注Notes: AAT. 年平均温度annual average temperature, VAAT. 年平均温度变异系数annual average temperature variation coefficient, AAPR. 年平均降水量annual average precipitation, VAAPR. 年平均降水量变异系数annual average precipitation variation coefficient, SS. 日照时数sunshine duration, VCSS. 日照时数变异系数sunshine variation coefficient, VCSP. 日照百分率变异系数sunshine percentage variation coefficient, VAAVR. 年平均水汽压变异系数annual average vapor pressure variation coefficient, AAP. 年平均气压annual average air pressure, VAAP. 年平均气压变异系数annual average air pressure variation coefficient; 下同the same below
使用COⅠ通用引物进行PCR扩增(北京六合华大基因),COⅠForward:5’-GGTCAACAAATCATAAAGATATTG;COⅠReverse:5’-TAAACTTCAGGGTGACCAAAAAATCA-3’(Folmeretal.,1994)。反应体系(25 μL):12.5 μL Taq MIX(0.625 U Taq酶,0.2 mmol·L-1dNTP,1.5 mmol·L-1MgCl2;Lot:A3701A,TaKaRa,中国大连);上下游引物各 0.15 μL(100 μmol·L-1)、1 μL DNA模板(80~100 ng),不足部分以ddH2O补充。反应条件:94 ℃预变性2 min;94 ℃变性30 s,42 ℃退火40 s,72 ℃延伸50 s,35个循环;72 ℃延伸5 min。PCR产物送北京六合华大基因科技股份有限公司进行纯化测序。
1.4 数据分析
双向测序的结果用ContingExpress进行拼接,通过CLUSTAL_X 2.0(Larkinetal.,2007)进行序列比对,并辅以人工校对。DnaSP 5(Librado & Rozas,2009)统计单倍型。ARLEQUIN 3.5(Excoffier & Lischer,2010)计算种群多样性指数,如单倍型多样度(h)、核苷酸多样度(π)、序列平均核苷酸差异数(k)、扩张系数(Tau)、种群分化系数(Fst);分子方差分析(AMOVA)。
通过SPASS 19.0(Coakes,2013)对遗传多样性、环境因子分别进行Shapiro-Wilk正态分布检验、单因素方差分析、Pearson相关性分析。
本研究选用最大似然法(maximum likelihood,ML)和贝叶斯法(Bayesian inference,BI)构建单倍型系统进化树,jModelTest(Darribaetal.,2012)和MrModelTest(Nylander,2004)中的赤池信息准则(Akaike information criterion,AIC)分别确立最适模型TVM+I+G。ML树和BI树构建均以琉球花棘石鳖Acanthopleuraloochooana和函馆锉石鳖Ischnochitonhakodaensis为外群。基于得到的最佳替换模型,使用PhyML(Guindonetal.,2010)构建ML树,采用启发式搜索,自举检验的数值为1 000,并计算各拓扑分值的支持率。使用MrBayes 3.2.5(Ronquistetal.,2012)构建BI树:采用马尔科夫链的蒙特卡罗方法(4个MCMC模拟)运算3×106,采样密度1/1 000。收敛性由分歧频率的平均标准偏差决定(<0.01),老化值分别设置为1.2×106代,剩余的代数用来构建50%一致树。最终得到一致的系统发生结构,而只在后验概率值上有轻微差异。结果使用TreeView(Surhoneetal.,2010)查看,通过ITOL在线工具(http://itol.embl.de/)进行系统树图像美化。基于中介网法(median-joining method,MJ)(Bandeltetal.,1999)和最大简约法(maximum parsimony,MP)(Polzin & Daneshmand,2003),通过Network 5.0.0.1绘制单倍型网络图。
2 结果与分析
2.1 遗传多样性
126只红条毛肤石鳖样本的mtDNACOⅠ基因部分序列长度为654 bp,碱基组成:A(20.51%)、T(41.72%)、C(14.23%)、G(23.54%),共发现41个多态位点,38个转换,4个颠换,无插入/缺失,界定29种单倍型,单倍型最多的采样点是长岛和大连(表2)。总体单倍型多样性0.899±0.013,核苷酸多样性0.013 3±0.006 8,核苷酸两两差异度8.689±4.038。单倍型多样性最高的为砣矶岛(0.955±0.057),最低的为葫芦岛(0.742±0.084);核苷酸多样性最高的为大连(0.013 9±0.007 5),最低的为秦皇岛(0.007 6±0.004 3)。红条毛肤石鳖各地理种群(除蓬莱外)的遗传多样性指数正态性检验结果表明,所有遗传多样性指数都服从正态分布;样本量之间的相关性分析结果显示,各多样性指数均不受个体数量的影响,说明可以用此数据表示各地理种群的遗传多样性。

表2 红条毛肤石鳖种群遗传多样性Table 2 Sampling information and diversity indices for 9 Acanthochiton rubrolineatus populations
注: inf为无限大, —为数据未收录, 括号内为标准差; 下同
Notes: inf. infinite, —. no data, standard deviation in brackets; the same below
2.2 环境因子对遗传多样性的影响
除砣矶岛(未被国家气象数据共享服务收录)外,所有环境因子经检验均为正态分布。通过Person相关性检验,发现红条毛肤石鳖种群核苷酸多样性与纬度、年平均温度变异系数呈显著负相关(rπ-LA=-0.808、rπ-VAAT=-0.795、rπ-AAP=-0.903、rh-AAP=-0.921;P<0.05),其他数据间无显著相关性(P>0.05)(表3)。

表3 红条毛肤石鳖遗传多样性与环境因子的相关性分析Table 3 Relationship correlations between genetic diversity of Acanthochiton rubrolineatus and environmental factors
注: LO. 东经, LA. 北纬; 下同
Notes: LO. east longitude, LA. northern latitude; the same below
2.3 遗传结构
基于COⅠ基因的两两种群之间的Fst值为-0.253(烟台-蓬莱)~0.565(秦皇岛-蓬莱)(表4)。秦皇岛与莱州、大连、长岛、砣矶岛、烟台、蓬莱6个种群之间存在显著的低水平遗传分化(0.121~0.565),葫芦岛与莱州、大连、长岛、烟台存在显著的低水平遗传分化(0.168~0.378),营口与莱州、长岛、烟台存在显著的低水平遗传分化(0.231、0.229、0.232),砣矶岛与烟台存在显著的低水平遗传分化(0.113)。
AMOVA表明,红条毛肤石鳖种群遗传变异主要发生在种群内(83.26%,P<0.001)(表5),较少部分的遗传变异来自于种群间(16.74%,P<0.001)。表明环渤海红条毛肤石鳖的遗传分化主要表现为种群内分化。

表4 基于COⅠ基因红条毛肤石鳖种群间遗传分化指数Table 4 Pairwise estimated values of genetic differentiation coefficient among natural populations of Acanthochiton rubrolineatus
注: 种群分化系数(对角线下方)与基因流(对角线上方);*P<0.05
Note:Fst(below diagonal), and gene flow (above diagonal);*P<0.05

表5 红条毛肤石鳖分子方差分析Table 5 AMOVA of Acanthochiton rubrolineatus
系统进化树(图2)显示,环渤海红条毛肤石鳖没有形成明显的进化分支。单倍型网络图(图3)与进化树一致。

图2 红条毛肤石鳖COⅠ基因的BI和ML系统发生树Fig. 2 Phylogenetic tree (BI and ML) based on COⅠ gene of Acanthochiton rubrolineatus
分支处数字代表后验概率/自举检验值
The number at the branch represents posterior probability/bootstrap test value
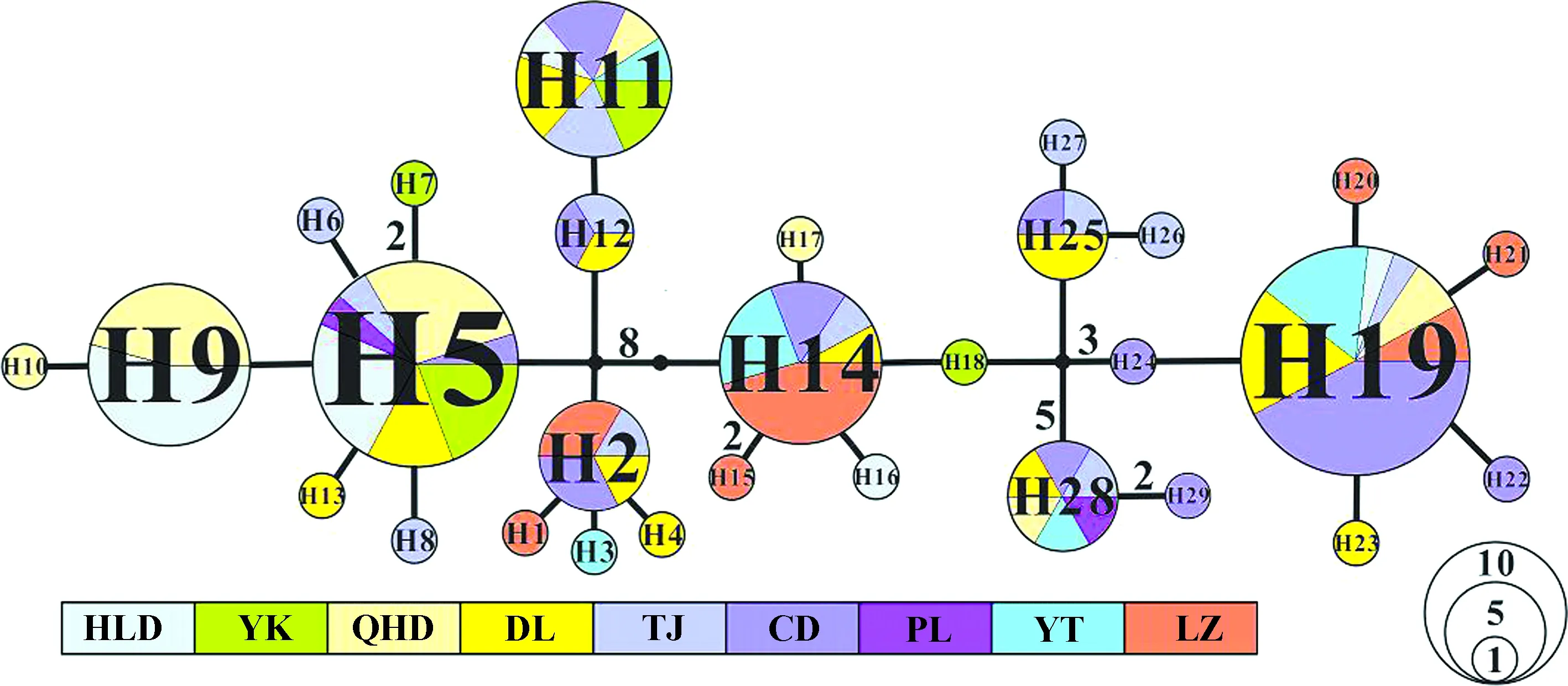
图3 基于COⅠ基因的红条毛肤石鳖单倍型网络Fig. 3 Haplotype network based on COⅠ gene of Acanthochiton rubrolineatus
2.4 种群历史动态
核苷酸错配分布分析表明,观察值呈近似单峰分布(图4),分布差异无统计学意义(SSD=0.017,P=0.220>0.05;Harpending’s raggedness index=0.027,P=0.080),符合种群扩张模型下的预期分布。根据石鳖类线粒体COⅠ基因进化速率(1.2%每百万年)计算,红条毛肤石鳖种群的扩张时间约为111.86万年。

图4 红条毛肤石鳖的错配分布图Fig.4 Mismatch distributions of Acanthochiton rubrolineatus
3 讨论
红条毛肤石鳖8个种群(除蓬莱外)的线粒体COⅠ基因序列研究表明,各地理种群具有较高的单倍型多样性(0.742~0.955)和较低的核苷酸多样性(0.007 6~0.013 9),符合Grant和Bowen(1998)提出的海洋类生物较高的单倍型多样性和较低的核苷酸多样性模式,且与其他海洋无脊椎动物类似(Nietal.,2012;Zhouetal.,2016;史博洋等,2017;王玲等,2018)。红条毛肤石鳖126只个体获得29种单倍型(23.02%),种群的遗传多样性水平较低。低水平遗传多样性的物种易受到环境及人为因素影响,在不良环境条件下生存会受到威胁,应重视环渤海红条毛肤石鳖种群保护。
核苷酸多样性是评价种群遗传多样性的重要指标。环渤海红条毛肤石鳖8个种群的核苷酸多样性与纬度呈显著负相关,环渤海南部种群遗传多样性高于北部种群,推测南部种群更为古老。核苷酸多样性与年平均温度变异系数呈显著负相关,表明环境温度越稳定,红条毛肤石鳖遗传多样性越高,与其他海洋无脊椎动物生态遗传研究结果基本一致(Monarietal.,2007;Yuetal.,2009;史博洋等,2017)。
海洋生物通常在其分布的空间范围内具有较低水平的遗传分化,无显著遗传结构的特点(王玲等,2018)。大多数海洋无脊椎动物具有浮游幼虫期,幼虫在海流的推动下扩散,不同地理种群间进行基因交流,呈较低水平的遗传分化,表现出遗传均质性(Palumbi,1994;Lee & Boulding,2010)。环渤海红条毛肤石鳖种群间的Fst值较低且多数不显著(63.89%),种群间没有形成显著的遗传分化。同时,环渤海红条毛肤石鳖没有形成显著的遗传谱系结构,遗传分化程度低,种群间存在基因交流。红条毛肤石鳖发育过程经历担轮幼虫期(张均龙等,2013),推测在渤海环流推动下,各地理种群间基因交流频繁,这可能是环渤海红条毛肤石鳖没有形成显著地理格局的原因。
环渤海红条毛肤石鳖经历了种群扩张,根据扩张时间,环渤海红条毛肤石鳖的扩张发生在早更新世晚期(第二温暖期),也是多瑙-恭兹间冰期发生时间。这一时期,海平面上升,红条毛肤石鳖从冰期避难所向外扩张,占据新的栖息地,从而形成新的遗传格局(Shenetal.,2011;Nietal.,2012),推测红条毛肤石鳖属于间冰期扩张物种。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
当代水产(2022年5期)2022-06-05
环境工程技术学报(2022年3期)2022-06-05
烟台果树(2021年2期)2021-07-21
艺术品鉴(2020年6期)2020-08-11
汽车维修与保养(2019年4期)2019-09-11
走向世界(2019年18期)2019-08-16
商情(2017年19期)2017-07-14
