
不同温度下草地贪夜蛾年龄阶段实验种群两性生命表的构建
2019-02-10 10:50谢殿杰张蕾程云霞江幸福
植物保护 2019年6期
谢殿杰 张蕾 程云霞 江幸福
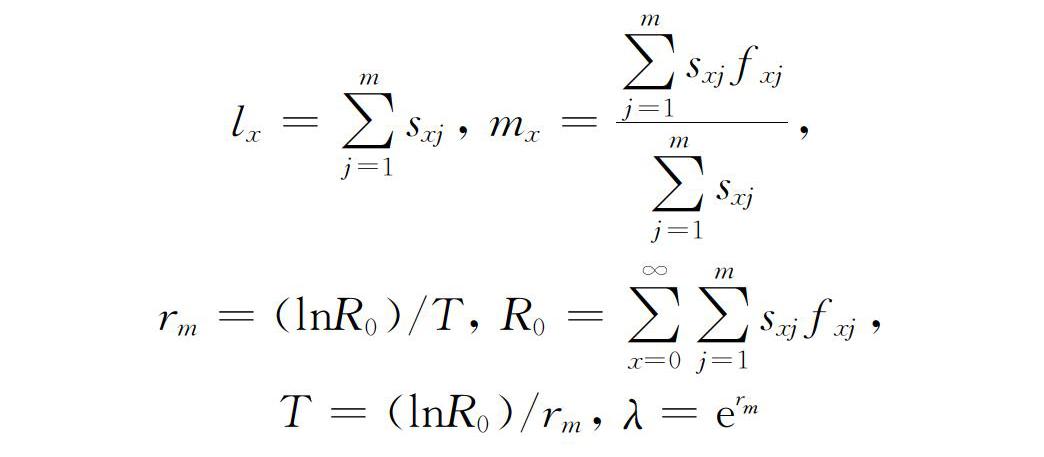


摘要 :為明确不同温度对草地贪夜蛾种群动态的影响,建立草地贪夜蛾种群动态模型,本研究通过建立年龄阶段两性种群生命表得到了室内不同温度下草地贪夜蛾种群的生命表参数,并以此为基础预测了未来2个月内草地贪夜蛾的种群动态。结果表明:不同温度(20、24、28、32、36℃)条件下草地贪夜蛾各阶段发育历期、存活率及繁殖力有显著差异。草地贪夜蛾的世代周期随温度降低而显著延长。20℃条件下世代周期最长,为(75.25±1.23)d;36℃条件下最短,为(24.79±0.36)d。草地贪夜蛾从卵存活到成虫的概率32℃下最高,为80%;20℃下最低,仅为24%。24℃条件下单雌平均产卵量为(1 448.83±97.64)粒,显著高于其他温度条件下。24℃条件下,种群净增长率(R0)最高,为420.16±71.48。内禀增长率(r)随温度升高先升高后降低,在28℃时达到最高值。根据种群生命表参数预测未来60 d内种群动态结果表明,较高温度有利于草地贪夜蛾种群世代数的增加,适当低温下产卵量大,更有利于种群数量的增加。本研究结果表明草地贪夜蛾有较广的适宜生存的温度范围,在20~36℃温度范围内均能完成生长发育和生殖。24~32℃是幼虫的最适生长发育温度范围,24℃是成虫最适的繁殖温度。
关键词 :草地贪夜蛾; 温度; 年龄阶段两性生命表
中图分类号:
S431.2
文献标识码: A
DOI: 10.16688/j.zwbh.2019485
Age stage two sex life table for laboratory populations of fall armyworm, Spodoptera frugiperda at different temperatures
XIE Dianjie, ZHANG Lei, CHENG Yunxia, JIANG Xingfu
(State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection,
Chinese Academy of Agricultural Sciences, Beijing 100193, China)
Abstract
In order to investigate the influence of different temperatures on Spodoptera frugiperda population dynamics and build the population dynamics model, the age stage two sex life table was used to obtain life table parameters at the different temperatures, and the population dynamics of S. frugiperda in the next two months were predicted. The results showed that different temperatures (20, 24, 28, 32, 36℃) had significant different effects on the duration of different developmental stage, survival rate and fecundity of S. frugiperda. The individuals reared at 20℃ and 36℃ had the longest (75.25 d±1.23 d) and the shortest (24.79 d±0.36 d) generation cycles, respectively. The pre adult surviving rates of the individuals reared at 32℃ was the highest (80%) and that at 20℃ was the lowest, only 24%. The average numbers of egg laid by single female adult reared at 24℃ was 1 448.83±97.64, which was significantly higher than those reared at other temperatures. The net reproductive value (R0) of individuals reared at 24℃ was the highest, 420.16±71.48. With the rise of temperature, the intrinsic growth rate of increase (r) increased first and then decreased, and reached the highest at 28℃. According to the parameters of life table, the prediction of population dynamics within next 60 days indicated that high temperature was beneficial to the increase of generation number, and high fecundity at low temperature was more conducive to the increase of population size. The results showed that the S. frugiperda has a wide range of suitable survival temperatures, and can complete the growth and reproduction within the temperature range of 20-36℃. The most suitable temperature range for larvae is 24-32℃, and the optimum reproduction temperature for adults is 24℃.
Key words
Spodoptera frugiperda; temperature; age stage two sex life table
草地贪夜蛾Spodoptera frugiperda (J. E. Smith)属鳞翅目Lepidoptera,夜蛾科Noctuidae,灰翅夜蛾属 Spodoptera,原产于美洲热带和亚热带地区[1]。草地贪夜蛾具有寄主范围广,为害严重,繁殖能力强,适应性强等特点,在中、南美洲可周年发生[23],成虫具有迁飞习性,在中、南美洲可向南迁飞至阿根廷,向北迁飞至加拿大[46]。近年来受到国际贸易和气候变化的影响,2016年1月,草地贪夜蛾首次入侵非洲,并迅速蔓延到非洲大部分地区[7],以及亚洲部分国家[8],成为一种世界性农业害虫。2019年1月草地贪夜蛾入侵我国云南,随后迅速蔓延,截至目前,已经有21个省(市)受到草地贪夜蛾为害[911]。
温度、湿度、光周期等环境因素影响着昆虫的发育历期、存活率、繁殖力及分布范围,其中,温度是最主要的环境因素[12]。昆虫种群的繁衍需要适宜的温度范围,过高或过低的温度都会对种群繁衍产生不利影响[13]。在20~32℃,甜菜夜蛾Spodoptera exigua的发育速率随着温度的升高而加快[14]。东方黏虫Mythimna separata在18~30℃都能完成生长发育和生殖,24℃下种群存活率高、繁殖力大、世代周期较短,更适合种群增长[15]。高温对烟粉虱Bemisia tabaci、谷象Sitophilus granarius的生长发育和繁殖有显著的抑制作用[1617]。
有关温度对草地贪夜蛾的影响国内已经有少量报道,如何莉梅等研究了草地贪夜蛾的发育起点温度和有效积温[18];鲁智慧等研究了6个温度(17、22、27、32、37、42℃)下草地贪夜蛾的存活率、发育历期、体重及繁殖力等[19],但均未计算种群生命表参数,也未进行种群的动态预测。关于不同温度对草地贪夜蛾种群动态的影响和两性生命表的研究尚未见报道。
生命表是种群生态学与害虫治理的重要工具,但是,传统生命表忽略了昆虫种群的世代重叠及个体间的发育差异[20],传统雌性生命表无法正确描述昆虫的变态且忽略雄性个体[21]。因此,近年来国内外学者普遍采用年龄阶段两性生命表对昆虫的阶段存活率、发育历期、繁殖力等进行研究。年龄阶段两性生命表已经广泛应用于橘小实蝇Bactrocera dorsalis[22]、白纹伊蚊Aedes albopictus和埃及伊蚊Aedes aegypti[23]、烟草甲Lasioderma serricorne[24]等多种昆虫的研究。本实验室已经利用年龄阶段两性生命表对3种黏虫近缘种,东方黏虫M. separata[15]、淡脉黏虫Mythimna roseilinea[25]、劳氏黏虫Mythimna loreyi[26]的生长发育和生殖进行了研究。
草地贪夜蛾在我国蔓延迅速,已经对我国的粮食安全产生了巨大威胁[2728],亟待建立完善的预测预报体系。温度作为其种群繁衍的决定因素之一,有必要进行深入探究。本试验通过设定不同饲养温度,构建了不同温度下草地贪夜蛾年龄阶段实验种群两性生命表,并对其种群动态进行了预测。确定其生长发育适宜的温度范围,为预测其可能的分布范围、不同地区的发生代数及该虫的预测预报和田间防治提供科学依据。
1 材料与方法
1.1 供试虫源
试验用草地贪夜蛾幼虫于2019年4月采自广西壮族自治区南宁市(108.37°E,22.82°N),在室内人工饲养繁殖,建立种群。饲养条件为:温度(28±1)℃、相对湿度60%±5%、光周期L∥D=16 h∥8 h、光照强度18 000 lx。幼虫使用新鲜幼嫩玉米苗饲养,饲养密度为10头/瓶(750 mL,高14 cm,直径8 cm),在含水量為15%的灭菌土中化蛹,成虫用10%蜂蜜水补充营养。
1.2 仪器设备
RXM 508C 恒温培养箱,宁波江南仪器厂;亚都超声波加湿器,北京亚都环保科技有限公司。
1.3 草地贪夜蛾室内实验种群的饲养及观察
试验设定20、24、28、32、36℃共5个温度梯度。试验均在恒温培养箱中进行,温差范围为±1℃。相对湿度为60%±5%,光周期为L∥D=16 h∥8 h、光照强度18 000 lx。取28℃条件下草地贪夜蛾同一天产的卵500粒,分为5组,100粒为1组装入750 mL玻璃瓶中,放入不同温度的培养箱。每天早(8:00)晚(20:00)各观察1次,记录卵孵化历期及存活情况。初孵幼虫转入45 mL指形管,每管中放入1头幼虫,用新鲜玉米叶饲喂。每天更换新鲜的玉米叶并记录各龄期的发育历期及幼虫存活情况。初孵幼虫到化蛹的时间为幼虫的发育历期。幼虫化蛹至羽化为成虫的时间为蛹期。成虫羽化当天,选取同天羽化雌雄成虫两两配对放入直径5 cm、高12 cm的透明塑料罩中,塑料罩口放置纱布供其产卵,用10%蜂蜜水补充营养。每天记录产卵量、存活情况等数据,直至所有成虫全部死亡。
1.4 年龄阶段两性种群生命表的构建及种群动态预测
利用年龄阶段两性种群生命表的原理统计原始数据[29],记录草地贪夜蛾各发育阶段的历期、幼虫存活率、化蛹率、羽化率、产卵量、雌雄成虫寿命等数据。并根据每日记录的产卵量和群体存活数计算年龄阶段特征存活率(sxj),即个体由卵发育到年龄x及龄期j时的概率;雌虫年龄阶段特征繁殖力(fx),即雌成虫在年龄x时的产卵量;种群年龄阶段特征存活率(lx),即从卵发育到年龄x的存活率;种群年龄阶段特征繁殖力(mx),即种群在年龄x时的平均产卵量;种群年龄阶段特征繁殖值(lxmx),即种群在年龄x时的净繁殖力。
种群生命表参数内禀增长率(rm)根据二分迭代法,用Euler Lotka公式计算[29];周限增长率为(λ),即在不受外界环境限制的情况下,种群平均每天增长的速率;净增长率(R0),即种群在一定条件下经过一个世代后的增殖倍数;平均世代周期(T)[30]按如下公式计算。
lx=∑mj=1sxj, mx=∑mj=1sxjfxj∑mj=1sxj,
rm=(lnR0)/T, R0=∑∞x=0∑mj=1sxjfxj,
T =(lnR0)/rm, λ=erm
1.5 草地贪夜蛾种群动态的预测方法
利用年龄阶段两性种群生命表软件TWOSEX MSChart[31]计算得到的数据,使用TIMING MSChart程序[32]预测5种温度条件下草地贪夜蛾未来2个月的种群动态情况。
1.6 数据分析
试验中所有数据的平均数和标准误用SPSS 20软件计算,对不同温度下草地贪夜蛾发育历期、产卵量、幼虫存活率、化蛹率、卵到成虫的概率进行方差分析,使用Tukeys HSD法进行差异显著性检验。利用TWOSEX MSChart和Origin 9.1绘图。
2 结果与分析
2.1 不同温度对草地贪夜蛾各虫态发育历期及繁殖的影响
草地贪夜蛾在20~36℃温度范围内均能完成整个世代,不同温度对草地贪夜蛾的生长发育及繁殖有显著影响。不同温度条件下草地贪夜蛾的平均世代周期存在显著差异。其中,在20℃条件下平均世代周期最长,为(75.25±1.23)d;36℃条件下最短,为(24.79±0.36)d(表1)。
草地贪夜蛾各虫态发育历期都随温度升高而显著缩短,36℃条件下最短,卵期(2.00±0.00)d,1~6龄幼虫期分别为(1.73±0.06)d、(1.12±0.12)d、(1.37±0.10)d、(2.00±0.10)d、(2.22±0.12)d、(2.78±0.10)d,蛹期(4.74±0.08)d。成虫寿命随温度升高显著降低,在24℃条件下显著高于其他温度,雌、雄蛾寿命分别为(17.00±0.88)d和(17.67±0.99)d。24℃條件下单雌平均产卵量为(1 448.83±97.64) 粒,显著高于其他温度条件下。20℃和28℃下的单雌平均产卵量显著高于32℃和36℃条件下(表1)。
2.2 温度对草地贪夜蛾种群存活率及繁殖力的影响
年龄阶段特征存活率(sxj)表示草地贪夜蛾从卵发育到年龄x及龄期j的概率。试验结果表明,不同温度条件下草地贪夜蛾的年龄阶段特征存活率存在显著差异。20~32℃条件下卵的孵化率达95%以上,其中32℃时卵孵化率为100%,36℃时卵的孵化率仅为75%。草地贪夜蛾从卵存活到成虫的概率在32℃下最高,为80%,20℃下最低,仅为24%。同时,温度对草地贪夜蛾雌雄成虫的比例也会产生显著影响,低温20℃和高温36℃条件下,雌蛾的数量显著高于雄蛾,32℃时,雄蛾数量显著高于雌蛾,而其他温度雌雄蛾数量基本一致(图1)。
年龄阶段特征繁殖力曲线由lx、mx、lxmx、fx共同组成。其中,lx表示不考虑阶段分化的种群年龄阶段特征存活率;mx表示种群在年龄x时的平均产卵量;lxmx为种群年龄阶段特征存活率lx与种群年龄阶段特征繁殖力mx的乘积;fx为雌虫年龄阶段特征繁殖力,表示雌成虫在年龄x时的产卵量。不同温度条件下,lx曲线趋势相同,即前期曲线较陡,后期趋于平缓,表明幼虫期是草地贪夜蛾个体死亡的主要阶段。36℃时繁殖力曲线出现得最早,为第21天;20℃时出现的最晚,为第61天。24℃时,草地贪夜蛾雌虫mx、lxmx和fx均高于其他温度,表明此温度是草地贪夜蛾较适宜的繁殖温度。此外,繁殖力曲线出现忽高忽低的现象,表明草地贪夜蛾羽化和产卵均不集中,且存在间断产卵的情况。
2.3 不同温度对草地贪夜蛾种群生命表参数的影响
草地贪夜蛾种群平均世代周期随温度升高而显著降低,20、24、28、32、36℃下依次为(67.86±1.40)d、(45.12±0.50)d、(30.24±0.69)d、(28.13±0.47)d、(24.09±0.34)d。24℃条件下,种群净增长率(R0)最高,为420.16±71.48,显著高于其他温度条件下。内禀增长率(r)随温度升高先升高后降低,在28℃时达到最高。一定范围内,温度升高可以提高种群的增长率,但是温度过高不利于种群的增长。24~36℃范围内,种群的周限增长率(λ)没有显著差异,20℃时周限增长率显著低于其他温度条件下。草地贪夜蛾种群周限增长率数据表明,在28℃条件下,如果其他环境因子适宜,草地贪夜蛾种群将会在1个世代周期(30.24 d)内以0.180的指数增长速率每天增长1.20倍。
2.4 不同温度下草地贪夜蛾种群动态预测
种群动态预测可为害虫综合防治提供理论依据。本研究利用100粒卵为起始虫数构建的年龄阶段两性生命表数据模拟预测了未来2个月内的草地贪夜蛾种群增长情况。结果表明,2个月后不同温度下草地贪夜蛾种群动态明显不同。此时,36℃下第三代大量幼虫进入蛹期,32℃下第三代处于幼虫期,28℃下第三代处于卵期,且二代蛹尚未完全羽化,24℃下第二代处于幼虫期,20℃下第一代处于蛹期,部分蛹开始羽化。同时,各温度条件下的种群数量也明显不同。32℃和28℃下第三代时幼虫数量超过5个数量单位;36℃下第三代时幼虫数量超过3个数量单位;24℃下第二代时幼虫数量超过3.5个数量单位,明显高于其他温度下第二代的幼虫数量。
3 讨论
昆虫对外界温度的变化非常敏感,在其生长发育的过程中,完成某一发育阶段所需的有效积温是固定的,故其发育历期会随温度变化而变化。温度对昆虫的生长发育、繁殖、行为、生活史等均有显著影响[3334]。本研究通过构建草地贪夜蛾的年龄阶段两性生命表,发现草地贪夜蛾的发育历期、存活率、繁殖力等种群参数在不同温度条件下差异显著。在一定温度范围内,草地贪夜蛾的发育历期随温度降低而显著延长,与其他昆虫一致[35]。20℃条件下草地贪夜蛾1个世代周期可长达75.25 d,36℃条件下最短,为24.79 d,与Schlemmer[36]、何莉梅等[18]的研究结果基本一致。与东方黏虫[15]相比,相同温度条件下草地贪夜蛾的世代历期更短,即相同时间内会产生更多世代,可能造成更严重的危害。低温20℃和高温36℃条件下,草地贪夜蛾从卵发育到成虫的比例显著降低,分别为24%和42%,且雌虫数量显著高于雄虫。20℃时,大量的草地贪夜蛾不能顺利完成羽化,在羽化过程中死亡。20、24、28、32℃和36℃条件下草地贪夜蛾的平均单雌产卵量分别为829.1、1 448.8、911.9、375.7粒和270.0粒,说明高温不利于其产卵,其最适产卵温度为24℃,且此温度的产卵前期显著短于其他温度。根据高温和低温条件下的世代周期、雌雄比例和产卵量,可以推测不利的外界环境会导致雌虫数目的增加,种群倾向于产生更多的后代来弥补不利环境对种群个体数量的影响。同时,我们可以明确草地贪夜蛾适宜温度范围广,在20~36℃范围内均能正常完成世代,最适生殖温度为24℃。我们前期研究表明,草地贪夜蛾的飞行适宜温度范围为24~32℃。最适飞行温度为32℃[37]。这说明,草地贪夜蛾生殖与飞行对温度的需求不同。
不同温度条件下,草地贪夜蛾的年龄阶段特征存活率及繁殖力明显不同。24~32℃下卵的孵化率及幼虫的存活率较高。内禀增长率、周限增长率和净增长率是重要的种群生命表参数,可反映种群增殖能力[38],可以用来预测种群未来的发展趋势。草地贪夜蛾在28℃下的内禀增长率和周限增长率最高,但是净增长率低于24℃条件下。因此,单一的生命参数不能准确衡量种群的发展潜能,需要对种群的生命表参数进行综合分析。本试验利用两性生命表的种群预测系统,综合多方面因素对不同温度下的草地贪夜蛾种群增长趋势进行了预测。结果显示,28、32、36℃下草地贪夜蛾种群世代数增长较快,但是增长量小于24℃下。
鲁智慧等研究了17、22、27、32、37、42℃下草地贪夜蛾不同阶段的存活率、发育历期、体重及繁殖力[19];何莉梅等测定了不同温度下草地贪夜蛾各阶段的发育起点温度和有效积温,并构建模型预测各阶段的发育速率[18],但都缺乏对生命表参数的分析及种群动态的预测。本试验研究了20、24、28、32、36℃下草地贪夜蛾的发育历期、存活率、繁殖力和种群生命表参数,并预测了不同温度下未来60 d的种群动态,对该虫的预测预报及防治策略的制定具有重大意义。当然,除温度外还有许多生物和非生物因素会影响昆虫的种群动态,如光周期对棉铃虫发育历期及蛹重有影响[39];高温和化学农药对西花蓟马F1代生命表参数有影响[40];不同寄主植物上草地螟种群动态有差异[41];湿度对草地螟发生为害规律有影响[42];不同豌豆品种对番茄尺蠖种群有影响[43];低氧胁迫对小菜蛾的两性生命表参数有影响[44]等。因此,由于多种因素的相互作用,在室内稳定条件下得出的草地贪夜蛾的种群动态与自然环境中的种群动态会存在一定差异,需要进一步进行田间试验,以更深入地了解野外环境下草地贪夜蛾种群的发生规律。草地贪夜蛾从2019年1月入侵我国以来已经扩散到除新疆、西藏、内蒙古、东北以外的大部分地區,明年其为害范围还将进一步扩大[10]。因此,急需对其种群动态的影响因素进行深入的研究,摸清其生长发育及繁殖规律,为虫情预测预报和虫害控制提供理论依据。
参考文献
[1] TODD E L, POOLE R W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the Western Hemisphere [J]. Annals of the Entomological Society of America, 1980, 73(6): 722738.
[2] MURU'A G, MOLINA OCHOA J, COVIELLA C. Population dynamics of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) and its parasitoids in northwestern Argentina [J]. Florida Entomologist, 2006, 89(2): 175183.
[3] LUGINBILL P. The fall army worm [R]. USDA Technology Bulletin, 1928, 34: 91.
[4] SPARKS A N. A review of the biology of the fall armyworm[J]. Florida Entomologist, 1979, 62(2): 8287.
[5] JOHNSON C G. Migration and dispersal of insects by flight[M]. London: Methuen, 1969: 745763.
[6] JOHNSON S J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the Western Hemisphere [J]. International Journal of Tropical Insect Science, 1987, 8(4/5/6): 543549.
[7] DAY R, ABRAHAMS P, BATEMAN M, et al. Fall armyworm: impacts and implications for Africa[J]. Outlooks on Pest Management, 2017, 28(5): 196201.
[8] 刘刚. 全国农技中心通报我国及周边国家草地贪夜蛾发生为害情况[J]. 农药市场信息, 2019(9): 59.
[9] 杨学礼, 刘永昌, 罗茗钟, 等. 云南省江城县首次发现迁入我国西南地区的草地贪夜蛾[J]. 云南农业, 2019(1): 72.
[10]吴秋琳, 姜玉英, 吴孔明. 草地贪夜蛾缅甸虫源迁入中国的路径分析[J]. 植物保护, 2019, 45(2): 16.
[11]郭井菲, 赵建周, 何康来, 等. 警惕危险性害虫草地贪夜蛾入侵中国[J]. 植物保护, 2018, 44(6): 110.
[12]BROUFAS G D, PAPPAS M L, KOVEOS D S. Effect of relative humidity on longevity, ovarian maturation, and egg production in the olive fruit fly (Diptera: Tephritidae)[J]. Annals of the Entomological Society of America, 2009, 102(1): 7075.
[13]FORSTER J, HIRST A G, WOODWARD G. Growth and development rates have different thermal responses [J]. The American Naturalist, 2011, 178(5): 668678.
[14]韩兰芝, 翟保平, 张孝羲. 不同温度下的甜菜夜蛾实验种群生命表研究[J]. 昆虫学报, 2003,46(2): 184189.
[15]秦建洋, 张蕾, 程云霞, 等. 不同温度下东方粘虫年龄阶段两性实验种群生命表的构建[J]. 植物保护学报, 2017, 44(5): 729736.
[16]GUO Jianying, CONG Lin, WAN Fanghao. Multiple generation effects of high temperature on the development and fecundity of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) biotype B [J]. Insect Science, 2013, 20(4): 541549.
[17]MOURIER H, POULSEN K P. Control of insects and mites in grain using a high temperature/short time (HTST) technique [J]. Journal of Stored Products Research, 2000, 36(3): 309318.
[18]何莉梅,葛世帅,陈玉超,等.草地贪夜蛾的发育起点温度、有效积温和发育历期预测模型[J].植物保护,2019,45(5):1826.
[19]鲁智慧,和淑琪,严乃胜, 等. 温度对草地贪夜蛾生长发育及繁殖的影响[J]. 植物保护,2019,45(5):2731.
[20]吴坤君, 龚佩瑜, 李秀珍, 等. 粘虫的年龄发育期结构生命表[J]. 昆虫学报, 1994,37(4): 426434.
[21]齐心, 傅建炜, 尤民生. 年龄龄期两性生命表及其在种群生态学与害虫综合治理中的应用[J]. 昆虫学报, 2019, 62(2): 255262.
[22]CHANG C, HUANG C Y, DAI S M, et al. Genetically engineered ricin suppresses Bactrocera dorsalis (Diptera: Tephritidae) based on demographic analysis of group reared life table[J]. Journal of Economic Entomology, 2016, 109(3): 987992.
[23]MAIMUSA H A, AHMAD A H, KASSIM N F A, et al. Age stage, two sex life table characteristics of Aedes albopictus and Aedes aegypti in Penang Island, Malaysia [J]. Journal of the American Mosquito Control Association, 2016, 32(1): 112.
[24]楊绍佳, 邱雯雯, 史梦竹, 等. 不同湿度下烟草甲实验种群两性生命表的构建[J]. 福建农业学报, 2018, 33(2): 177185.
[25]QIN Jianyang, LIU Yueqiu, ZHANG Lei, et al. Effects of temperatures on the development and reproduction of the armyworm, Mythimna roseilinea: Analysis using an age stage, two sex life table [J]. Journal of Integrative Agriculture, 2018, 17(7): 15061515.
[26]QIN Jianyang, ZHANG Lei, LIU Yueqiu, et al. Population projection and development of the Mythimna loreyi (Lepidoptera: Noctuidae) as affected by temperature: application of an age stage, two sex life table [J]. Journal of Economic Entomology, 2017, 110(4): 15831591.
[27]葛世帅, 何莉梅, 和伟, 等. 草地贪夜蛾的飞行能力测定[J]. 植物保护,2019,45(4):2833.
[28]郭井菲, 何康来, 王振营. 草地贪夜蛾的生物学特性、发展趋势及防控对策[J]. 应用昆虫学报, 2019, 56(3): 361369.
[29]CHI H, LIU H. Two new methods for the study of insect population ecology [J]. Bulletin of the Institute of Zoology, Academia Sinica, 1985, 24(2): 225240.
[30]YANG Yuting, LI Wenxiang, XIE Wen, et al. Development of Bradysia odoriphaga (Diptera: Sciaridae) as affected by humidity: an age stage, two sex, life table study [J]. Applied Entomology and Zoology, 2015, 50(1): 310.
[31]CHI H. TWOSEX MSChart: a computer program for the age stage, two sex life table analysis [OL]. http:∥140.120.197.173/Ecology/Download/TWOSEX.zip
[32]CHI H. TIMING MSChart: a computer program for the agestage, two sex life table analysis [OL]. http:∥140.120.197.173/Ecology/Download/TIMING.zip
[33]HOFFMANN A A, SRENSEN J G, LOESCHCKE V. Adaptation of Drosophila to temperature extremes: bringing together quantitative and molecular approaches[J]. Journal of Thermal Biology, 2003, 28(3): 175216.
[34]CUI Xuhong, WAN Fanghao, XIE Ming, et al. Effects of heat shock on survival and reproduction of two whitefly species, Trialeurodes vaporariorum and Bemisia tabaci biotype B[J]. Journal of Insect Science, 2008, 8(1): 24.
[35]徐盼, 徐志宏, 李绍进, 等. 不同温度下康氏粉蚧实验种群生命表[J]. 昆虫学报, 2012, 55(12): 13621367.
[36]SCHLEMMER M. Effect of temperature on development and reproduction of Spodoptera frugiperda (Lepidoptera: Noctuidae)[D]. Evanston: North West University, 2018.
[37]謝殿杰, 张蕾, 程云霞, 等. 温度对草地贪夜蛾飞行能力的影响[J].植物保护,2019,45(5):1317.
[38]FARHADI R, ALLAHYARI H, CHI H. Life table and predation capacity of Hippodamia variegata (Coleoptera: Coccinellidae) feeding on Aphis fabae (Hemiptera: Aphididae)[J]. Biological Control, 2011, 59(2): 8389.
[39]陈元生, 段德康, 陈超, 等. 光周期和温度对棉铃虫发育历期及蛹重的影响[J]. 环境昆虫学报, 2012, 34 (4): 407414.
[40]左太强,张彬,张绍婷,等.高温和啶虫脒处理西花蓟马对其F-1代生命表参数的联合作用[J].昆虫学报,2015,58(7):767775.
[41]张同强, 程云霞, 张蕾, 等. 用年龄阶段两性种群生命表研究不同寄主上草地螟种群动态的差异[J/OL]. 植物保护: 18[20190912].https:∥doi.org/10.16688/j.zwbh.2019013.
[42]罗礼智, 程云霞, 唐继洪, 等. 温湿度是影响草地螟发生为害规律的关键因子[J]. 植物保护, 2016, 42(4): 18.
[43]ALAMI S, NASERI B, GOLIZADEH A, et al. Age stage, two sex life table of the tomato looper, Chrysodeixis chalcites (Lepidoptera: Noctuidae), on different bean cultivars [J]. Arthropod Plant Interactions, 2014, 8(5): 475484.
[44]刘天生, 温梅芳, 柯富士, 等. 低氧胁迫下小菜蛾的两性生命表研究[J]. 福建农业学报, 2017, 32(3): 305311.
(责任编辑:杨明丽)
猜你喜欢
农家科技(2019年7期)2019-10-08
农家科技(2019年7期)2019-10-08
农家科技(2019年8期)2019-09-19
智族GQ(2019年8期)2019-08-30
南方农业·下旬(2019年4期)2019-08-26
山东农业科学(2019年6期)2019-08-16
现代农业科技(2019年12期)2019-08-07
小天使·二年级语数英综合(2018年7期)2018-09-11
文学港(2018年1期)2018-01-25
知识就是力量(2016年9期)2016-11-02
