
可用于二氧化碳捕获过程的微生物碳酸酐酶的挖掘与改造
2019-01-30 08:19蔡丽希楚云猛张光亚
生物工程学报 2019年1期
蔡丽希,楚云猛,张光亚
华侨大学 化工学院 生物工程与技术系,福建 厦门 361021
自工业革命以来,人类主要依赖应用煤炭、石油与天然气等化石燃料,导致大气中CO2的浓度急剧上升,加重地球表面的温室效应,引起全球气候变暖,海平面不断上升和病虫害增多等现象[1-3]。火力发电是目前最大的人为CO2排放源[4]。到2014年为止,全球火力发电比例仍然占了总供电量的66.7%,而且短期内这一比重并不会有所变化。在这样一个高度依赖化石燃料发电模式的大环境下,碳捕获和封存技术 (Carbon capture and storage,CCS) 在近些年来一直是碳减排等研究领域中的一个备受关注的热点问题[2,5-6]。
碳捕获和封存技术指的是将CO2从火力发电厂、钢铁厂、化工厂等排放源头分离出来,输送到封存地点并与大气长期隔绝的过程。主要分为捕集、运输、封存和监测四部分[7]。燃烧前CO2捕集技术 (PCC),特别是化学胺法捕集CO2技术是目前公认的最适合于捕集火电厂烟气中CO2的手段。CO2捕集一般常见于碱性溶液的吸收和随后的高温解吸塔解吸并压缩和封存,然而此过程面临一些技术挑战,首要问题是CO2解吸过程中高热能的损失[8]。可仿生矿化CO2的碳酸酐酶具有解决热能损失的潜力,嗜热又耐碱的碳酸酐酶可以加速CO2在高温化学吸收剂的吸收速率,并催化CO2转化为HCO3-,最终模拟地质学过程并生物矿化形成CaCO3沉淀,其终产物在整个地质学周期内性质非常稳定,对环境友好,因此利用碳酸酐酶的生物矿化CO2法是目前最具前景的CO2减少排放技术手段之一[9]。本文先简要介绍了碳酸酐酶,再从碳酸酐酶的挖掘、改造及固定化3个方面综述了国内外最新研究进展。
1 碳酸酐酶简介
碳酸酐酶(Carbonic anhydrases,CA,EC4.2.1.1)是一类能高效催化二氧化碳与水和碳酸与氢质子之间的可逆反应的锌金属酶,pH值介于4–9之间,且在温度低于65 ℃的条件下可以保持较高的活性和稳定性[10]。它是1933年由Meldrum和Roughton从牛红细胞中发现的第一个锌金属酶。在人类和动物的血液中,碳酸酐酶是红细胞中含量与功能重要性仅次于血红蛋白的最主要的蛋白质组分。碳酸酐酶是目前为止反应速率最快的几种酶之一,它的反应速率受到底物的扩散速率限制。不同家族的碳酸酐酶的催化速率为104–106/s[11]。CA在真核生物体中几乎是无所不在的,主要在生物体内起二氧化碳浓缩、光合作用、离子交换、呼吸作用以及pH稳定等作用。
到目前为止,已发现碳酸酐酶有α、β、γ、δ、ζ、η等6个不同家族,每个家族之间的氨基酸序列无明显同源性,是趋同进化的典型代表[7]。六大家族的碳酸酐酶的特点见表1,α-CA常见于哺乳动物、原生动物、藻类、植物、细菌、真菌、古菌的胞浆中,所有动物来源的碳酸酐酶都出自α-CA,是现今研究最透彻的家族,人体中含有分布于多种器官中的α-碳酸酐酶,其中催化活性最高且研究最为透彻的为人碳酸酐酶Ⅱ(hCAⅡ);β-CA首先发现在高等植物的叶绿体中,随后又在细菌、真菌、藻类、古菌中找到,在植物光合作用中,主要起 CO2固定及维持CO2浓度等作用;γ-CA则主要存在于一些细菌和古菌中;δ-CA 与ζ-CA主要存在于海洋的硅藻中;η-CA是近年来才发现的类型,主要存在于疟原虫中。这六大家族的酶结构虽然相似度不高,但每种酶的活性中心都会结合一个对催化过程起着重要作用的金属离子。其中加快CO2水合的核心机理就是金属离子对CO2的亲核攻击[9]。
碳酸酐酶生物矿化本质是催化Ca2+与水中的CO2之间发生的反应[7]。反应历程可以用如下方程式描述:
1) 气态的CO2溶于水,形成松散的水化CO2状态:

2) 在碱性条件下,CO2与水反应生成H2CO3:

3) H2CO3电离为质子和 HCO3-:

4) 在碱性条件下,HCO3-进一步电离形成CO32-和H2O:

5) 在Ca2+存在的情况下,生成CaCO3沉淀:
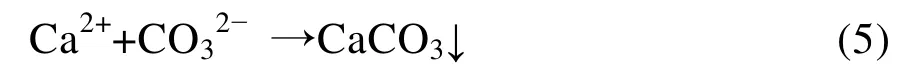
该反应的限速步骤为第2步CO2与水反应生成H2CO3。在自然界,CO2的水合反应速率相当缓慢[16-17],其转化数仅为1.3×10–1/s[18],当碳酸酐酶催化上述反应时,CO2水合转化数显著提高至1.4×107/s,约是自然界的107倍。但CA矿化过程中极度依赖反应液pH值,当pH<8时,反应2中CA活性明显降低,培养液中无机碳主要以CO2形式存在[19]。当pH>8时,由于反应液中含有OH-,反应2速率加快,HCO3-与CO32-不断生成促成CaCO3沉淀。
因此,微生物在仿生生成碳酸盐的过程中需要在碱性条件下进行,碳酸酐酶在pH值大于8的情况下才能保持较高的活性。而火电厂排出的烟气温度较高,接近60 ℃,一般的碳酸酐酶在高热与高碱下稳定性与活力较差,这极大地限制了碳酸酐酶的应用。有3种方法解决这个问题:1)从极端微生物(环境)中筛选出能够在温度高于50 ℃且pH>8条件下稳定发挥功能的碳酸酐酶;2) 使用蛋白质工程技术改造现有酶,使它变成热、碱稳定性酶[20],3) 通过固定化技术提高酶稳定性及增加重复使用性,降低成本。
2 微生物来源的嗜热、耐碱碳酸酐酶
近年来,极端微生物来源的碳酸酐酶因其独特的优势引起了广泛关注。极端微生物主要来源于深海、上下对流层、火山、矿井等极端环境,地球上的极端环境孕育了丰富的极端微生生物资源,这成为嗜热耐碱碳酸酐酶的重要来源。目前文献报道的已筛选到的嗜热、耐碱碳酸酐酶及其酶学性质见表2。其中几个比较有趣的碳酸酐酶描述如下。

表1 六大家族碳酸酐酶的性质与特点Table 1 Properties and characteristics of the six classes of carbonic anhydrases
2012年分离自美国黄石公园温泉的Sulfurihydrogenibium yellowstonenseYO3AOP1的碳酸酐酶 (SspCA) 是热稳定性最高的几种酶之一[21],以CO2为底物时,当温度为20 ℃、pH为7.5时,酶的动力学参数为kcat=9.35×105/s;kcat/Km=1.1×108/((mol/L)·s),同时SspCA具有酯酶活性,最适pH为9.6,最适温度为95 ℃[22]。酶的热稳定性实验表明SspCA在40 ℃与70 ℃的半衰期分别为53 d与8 d。SspCA的晶体结构显示了该酶高热稳定性的机理,如图1A所示,SspCA具有α-CA典型的二聚体结构,每个单体含有一个三角锥活性中心,由Zn2+与His89、His91、His108组成畸变四面体,其中使二氧化碳水化的机理核心便是Zn2+对于CO2的亲核攻击。乙磺酰唑胺 (AZM) 为活性中心共价结合的抵制剂。SspCA组成一个具有大界面区域特征的二聚体。主要由周围环绕着α-螺旋和小β-折叠的10个平行的β-折叠组成,活性中心位于从蛋白质表面至中心的一个圆锥体腔里,其蛋白质表面含有较多的带电氨基酸,形成一个较复杂的离子网络,这就是为什么相比与其他常温CA其热稳定性更高的原因。
从来自亚速尔群岛温泉极端微生物Sulfurihydrogenibium azorense中分离的碳酸酐酶(SazCA)是世界上现今为止催化活力最高的,它是典型的α-CA。以CO2为底物时,在20 ℃、pH为7.5条件下,酶动力学参数分别为kcat=4.40×106/s–1;kcat/Km=3.5×108/((mol/L)·s),最适pH为9.6,可以在0–100 ℃范围内保持活力。阴离子抑制实验表明HCO3-、CO32-、SO42-均对酶无明显的抑制作用。SazCA既嗜热又耐碱,酶活力又高,若能进一步提高其热稳定性,将是一个极具潜力的可应用于工业CO2捕集的碳酸酐酶[22-23]。
2015年,Simone课题组[38]揭示了SazCA的晶体结构,丰富了来自极端微生物的α-CA的结构信息。如图1B所示,SazCA的单体结构主要是由10个平行的β-折叠组成,周围环绕着3个α-螺旋、310-螺旋和小β-折叠。由Cys24与Cys178所组成的二硫键在不同来源的α-CA中相对保守。SazCA与SspCA的结构具有61.3%的相似度,SspCA上的Glu2和Gln207残基被SazCA的His2和His207取代,这种取代可能影响了His64的pKa值进而增强其质子穿梭能力,这些结构信息为设计高催化活性的α-CA提供了诱人的前景。
Littlechild课题组报道了一个现今催化活性居第二的碳酸酐酶(TaCA),该酶源于Thermovibrio ammonificans,热稳定性较高[24]。当温度为25 ℃、pH为7.5时,酶的动力学参数为kcat=1.6×106s–1;kcat/Km=1.6×/((mol/L)·s),TaCA在40 ℃与60 ℃的半衰期分别为152 d与77 d。而且在保温60 d后,酶在40 ℃与60 ℃的残余活性分别为91%与62%。与其他α-CA不同,TaCA具有特殊的四聚体结构,由2个与SspCA和SazCA结构相似的二聚体以二硫键连接形成,值得提及的是,形成分子间二硫键的2个保守的Cys残基Cys202和Cys47在SspCA和SazCA中未出现,四聚体结构可能是其保持高热稳定性的原因[24]。分析这些热稳定性CA的结构,一方面可以了解它们高热稳定性的分子机理,同时也为碳酸酐酶的改造提供了重要信息和方向。
上述基于微生物培养筛选的嗜热、耐碱的碳酸酐酶取得了长足进步,但从环境样品中 (宏基因组) 获取该酶的研究非常匮乏。最近,本课题组从几个深海热液口样品的宏基因组中筛选到16个疑似碳酸酐酶的基因,序列比对结果显示,其中1个与目前数据库中实验验证的碳酸酐酶相似度为46.9%,可能是一些新的嗜热碳酸酐酶并命名为VaCA。通过在线服务器SWISS-MODEL以Streptococcus mutans(PDBID:3LAS) 为模板同源建模,发现VaCA是一个相比TaCA结构较小但相对致密的四聚体结构,每个单体由1个类似右手的5个相互平行的β-折叠组成,周围环绕着3个α-螺旋和1个310-螺旋,分子内没有二硫键连接。目前正在进行相关的实验验证。从宏基因组中获取CA将进一步拓展其来源,为挖掘性能更佳的CA提供了可能。

表2 嗜热、耐碱的微生物碳酸酐酶的特性Table 2 Charactoristic features of the thermostable and alkalible of microbial carbonic anhydraes

图1 四种典型的极端碳酸酐酶的四级结构 (蓝色为锌离子,红色为抑制剂AZM)。(A) SspCA的二聚体结构,一个单体以黄色表示,另一个单体以绿色表示。(B) SazCA的二聚体结构。(C) TaCA的四聚体结构,四个单体分别以洋红色、绿色、蓝绿色和黄色表示。(D) VaCA的四聚体结构。Fig.1 The four typical quaternary structure of carbonic anhydrase.The zinc ion display in blue, and the inhibitor AZM are also shown in red.(A) The dimer structure of SspCA with one monomer shown in yellow, the other one in green.(B) The dimer structure of SazCA.(C) The tetramer structure of TaCA with four monomers shown in magenta, green,cyan and yellow respectively.(D)The tetramer structure of VaCA with four monomers.
3 蛋白质工程技术
尽管目前已经从自然界筛选到了大量的新型碳酸酐酶,但仍不能有效地满足环境应用与工业化生产中的迫切需求。蛋白质工程技术的发展,让研究者可以按照预期来调整并设计蛋白质的结构与功能[39]。蛋白质工程大致可分为定向进化和理性设计两大类。定向进化无需事先了解蛋白质的结构和功能,在实验室条件下模拟自然进化过程,通过随机突变和定向筛选获得更加适应环境压力的新的蛋白质[40-41]。Newman等[42]通过定向进化技术获得的碳酸酐酶突变体在工业条件下的CO2捕获能力提高了25倍,热稳定性提高了1000万倍,突变体可在50 ℃、4.2 mol/L浓度下的N-甲基-二烷酰胺 (MDEA) 下保持稳定,在75 ℃ 和4.2 mol/L MDEA 下半衰期为 20 h。这种策略可用来获得嗜热又耐碱的碳酸酐酶的高效催化酶。Alvizo等[43]对来自Deulfovibrio vuglaris中的β-CA进行定向进化来增加热稳定性,经过10轮的突变与筛选获得的碳酸酐酶可耐受107 ℃、pH大于10.0和4.2 mol/L浓度下MDEA等严苛环境,热稳定性与碱耐受性相比野生型酶提高了400万倍,同时CO2水合酶的能力也提高了24倍。这个热稳定性CA可以在高温 (87 ℃) 下用于CO2的生物矿化。Protelica 公司启动使用定向进化改造的碳酸酐酶用于碳捕获与封存这个项目(http://www.protelica.com/Carbonicanhydrase),旨在挖掘可耐受严苛的工业环境下的嗜热耐碱型碳酸酐酶。
理性设计基于己知蛋白质结构和功能的机理,并应用它们对突变位点进行设计,最终获得的突变体具有所预期的功能[44]。有研究者把常温CA改造成嗜热嗜碱型酶并获得成功。美国专利[45](专利号:7521217)调整人碳酸酐酶Ⅱ(hCAⅡ)分子表面的氨基酸,分别使Ala65、Phe93、Gln136、Lys153、Leu198、Ala247各自突变成Thr、Leu、His/Tyr、Asn、Met和Thr。使酶突变为极端嗜热酶,能在90 ℃稳定保持活性24 h。二氧化碳解决方案公司 (CO2Solutions Inc.) 将这个嗜热型碳酸酐酶投放至洗涤器的预分离高温烟气中,其CO2吸收率是以往技术的40倍 (http://www.CO2solutions.com/en/the-process)。Fisher等[46]在保证人碳酸酐酶Ⅱ (hCAⅡ) 原有的高酶活性和溶解性的情况下突变远离活性中心的表面氨基酸,将分子表面的氨基酸Tyr7、Leu224、Leu100、Leu240、Asn67和Asn62各自突变为Phe、Ser、His、Pro、Glu和Leu。使疏水性氨基酸突变成亲水性氨基酸来增加热稳定性,突变体融解温度上升 6 ℃。Kanth等[18]将Persephonella marinaEX-H1的碳酸酐酶 (PmCA) 的信号肽去除后在大肠杆菌表达,获得的无信号肽的同源二聚体PmCA (sp–) 其碱耐受性增强,热稳定性也大幅提高,在100 ℃中其半衰期为88 min,融解温度为84.5 ℃。Luca等[47]以现今发现催化活性很高的碳酸酐酶SazCA为模本,在其N端引入热稳定性最高的SspCA中所独有的6个His,准备设计1个超级碳酸酐酶SupCA。然而,SupCA的催化活性与热稳定性比原有两种酶均低,这可能是由于过多His的引入破坏了活性中心周围的氢键网络,SupCA的设想虽然失败,但也为后继酶设计提供一定的参考。Jo等[25]利用来源于淋病奈瑟氏菌Neisseria gonorrhoeae中的NgCA开展全细菌细胞催化分析,从E.coli中筛选,全细胞催化在低pH下稳定,即使在pH值小于HCO3-与CO32-的pKa下,全细胞催化系统可以有效地矿化CO2。因此,可以降低用于维持高pH值的费用。应用以上策略调整CA的结构可以有效地捕获CO2,但是改造成功并获得应用的例子较少。
在蛋白质工程改造探索过程中,也有研究者们从不同角度与层次分别对CA的热稳定性机制进行了颇有价值的研究和探索。文中通过对现有研究结果进行总结和分析,得出以下可能因素:1) 二硫键可稳定CA的结构,降低解旋状态下CA的构象熵以维持其热稳定性[57]。二硫键能够维持CA的热稳定性机制已通过实验获得验证,如Jo等[48]在NgCA中成功引入二硫键后,使得该酶的融解温度较引入二硫键之前提高了7.8 ℃,热稳定性也得到相应的提高。2) CA热稳定性主要取决于其分子构象的刚性程度,生物信息学揭示酶的刚性越强其热稳定性越强,而在CA表面的连接α-螺旋与β-折叠的loop圈具有很大的柔性,蛋白质的稳定性可以通过添加脯氨酸残基提高表面loop圈的刚性,或者通过减少loop圈的方式来增强蛋白质的表面致密性,因此,Boone等[49]用脯氨酸取代hCAⅡ表面loop圈的Glu234,设计出新颖的hCAⅡ突变体,热稳定性分析表明该突变体的融解温度相比野生型提高了3 ℃。另一方面,包含230–240个氨基酸残基的区域形成了一个舒展的loop圈,具有特殊的热不稳定特征。删除后其热稳定性增高,这一影响因素在热稳定性高的SspCA中得到了进一步的支持。其蛋白质的表面致密性较高,且无此loop圈。这一发现对改善野生型酶的融解温度具有一定的指导意义。3)温度B因子 (B-factor) 的影响,B-factor是理性改造酶热稳定性的一个重要指标,氨基酸的B-factor值越大,它的柔性就越大,不利于CA的热稳定性,因此Jo等[48]利用从头设计的方法在NgCA中计算出温度B因子 (B-factor) 与氨基酸残基对柔韧度大于25的3对氨基酸残基,各自突变成3对Cys,成功引入二硫键并形成3个突变体,其中1个突变体的热动力学与热稳定性都大幅增强,这可能归功于B-factor与二硫键的引入。
4 碳酸酐酶的固定化
除了通过筛选和改造获得嗜热、耐碱的碳酸酐酶外,对CA进行固定化也可以一定程度上达到上述目的。而且,游离酶在强酸碱、高温、高压、有机溶剂等条件下易失活,且由于直径过小(接近于50 Å),很难从吸收塔中回收,其工业化应用被大大地限制了。但固定化技术不仅有利于酶稳定性的提高,也有助于酶的分离,且固定化酶具有不溶性和一定的机械强度,可循环应用于反应器中[50-51]。
近年来,采用固定化CA 仿生法来捕集 CO2的技术成为了研究热点,国内外众多学者对固定化材料、固定化酶的制备方法和CO2的矿化效率展开了研究,为常温CA在工业上的应用提供了更具潜力的方法[52]。Prabhu等[53]将来源于短小芽孢杆菌Bacillus pumilus常温CA吸附在壳聚糖的表面,固定化酶对热和抑制剂作用的稳定性均有所提高,在50 ℃同等条件保温90 d后,固定化酶可保持60%的初始酶活,而游离酶只有30%,同时在5 min内CaCO3沉淀生成速率是游离酶的2倍。Ozdemir等[54]将来源于牛的碳酸酐酶 (BCA)固定在聚氨酯 (PU) 泡沫上,固定化酶相比游离酶的动力学参数有所下降,Km由12.2 mmol/L下降至9.6 mmol/L,但固定化酶的热稳定性明显增强,在反应器中连续工作7个循环后或在室温贮存45 d后,还可保持100%的酶活性,而游离酶即使在4 ℃贮存45 d后完全失活。Vinoba等[55]将BCA固定到经化学处理的磁性微球上,该固定化酶在反应器工作30个循环后,其CO2捕获能力是游离酶的26倍。且固定化酶在室温贮存30 d后,仍能保持82% 的初始酶活性。Yadav等[56]将BCA包埋于3种不同粒径海藻酸钠微球上,结果显示:粒径最小的微球上酶活性最高,相比游离酶,固定化酶热稳定性显著增强,其最适温度提高了近10 ℃,固定化酶在工作42 d后仍能保持67%的初始酶活。Capasso等[57]将SspCA固定在聚氨酯(PU) 泡沫上,其热稳定大大增强,可以在100 ℃下存活48 h,在70 ℃下长期工作,并可用于火电厂高温捕获CO2的反应器中。酶在载体上的固定化往往会让酶发生一定程度的构象变化,与底物的接触效率降低,促使催化活性降低。科学家发现纳米颗粒可以克服以上问题。Shanbhag等[58]将BCA溶解于自组装肽P114中形成50–200 nm的纳米颗粒,粒径越小,酶越不容易发生构象变化,该固定化酶可保持98%水化酶活性,并在低温 ((22±2) ℃) 和高温 (50 ℃) 下保持自组装状态。近年来,基于R5介导的二氧化硅纳米球的仿生制备原理,国内外学者做了二氧化硅为基质的酶自固定化研 究[59]。Jo等[60]成功地将R5和目标酶基因融合,经表达和纯化后,由 R5介导的二氧化硅纳米球自固定化酶,固定化后的碳酸酐酶可在50 ℃贮存5 d后仍保持80%的活性,且在60 ℃时酶活性是25 ℃的11倍,这有利于应用于后继工业化生产。
我们课题组将CA和类弹性蛋白多肽(Elastin-like polypeptides,ELPs) 融合表达,借助ELPs自分离技术经离心纯化目标酶,通过ELPs独特的仿生硅化能力,在形成二氧化硅粒子的同时实现CA的自固定化,固定化酶的最适温度较游离酶提高了10 ℃,可达80 ℃,其热稳定性大幅提高,但酶催化效率略有下降,酶活回收率为64.9%,固定化过程工艺参数仍有较大的优化空间。制备的固定化碳酸酐酶在酶浓度达到9 mg/mL时,封装率达到91%,在4 ℃放置72 h后,泄漏的酶蛋白仅为0.96%。由于ELPs介导的仿生硅化现象尚未见报道,其介导仿生硅化的分子机制及所得载体对酶活性影响规律仍有待深入,本课题组也将会持续探究。
5 展望
综上所述,在CO2浓度居高不下、全球温室效应加剧,但各项碳捕集技术尚不完善且成本较高的今天,碳酸酐酶作为一种新型的高效生物催化剂应用于CCS的前景格外引人注目。而极端微生物的挖掘、蛋白质工程技术和酶固定化技术的发展又恰恰为碳酸酐酶的工业化应用提供了诱人的前景。然而,当前大数据显示地球极端生境中还存在大量的CA基因等着被发现。这些大多是不可培养的微生物。宏基因组学与生物信息学工具在基因识别方面具备独特能力,一旦识别出新型的CA编码基因,就容易在不同的微生物宿主中克隆并表达,从而研究它们的性质及在二氧化碳生物矿化中的潜在用途。目前,尚未发现的数据很多,甚至有可能存在目前发现的6个家族之外的新的CA。与此同时,碳酸酐酶固定化可实现其重复利用,提高了操作稳定性,降低了运行成本。但是传统的固定化方法往往会导致酶活力降低或催化性能的下降。因此,拓展搜寻碳酸酐酶的范围,改良蛋白质工程改造技术,研发廉价、高效的固定化方法仍是后续研究的重点,这些技术和方法的创新将为减轻温室效应、延缓全球变暖这一迫切需解决的问题提供新思路。
猜你喜欢
食品科学技术学报(2022年5期)2022-10-11
肝博士(2022年3期)2022-06-30
海外星云(2021年9期)2021-10-14
分析化学(2019年4期)2019-05-13
陶瓷学报(2019年5期)2019-01-12
中国饲料(2016年17期)2016-12-01
中国塑料(2016年7期)2016-04-16
中国老年学杂志(2015年5期)2015-12-31
中国塑料(2015年6期)2015-11-13
中国塑料(2015年8期)2015-10-14
