
干扰SIRT7抑制甲状腺癌细胞的增殖、迁移和侵袭能力
2019-01-28 01:55陈晓曦陈旭旭林佳浩陈吉彩黄国裕
温州医科大学学报 2019年1期
陈晓曦,陈旭旭,林佳浩,陈吉彩,黄国裕
(1.温州医科大学附属第一医院 疝与腹壁外科,浙江 温州 325015;2.温州医科大学 第一临床医学院, 浙江 温州 325035;3.温州医科大学附属第一医院 胃肠外科,浙江 温州 325015)
Sirtuin(SIRT)家族(包括SIRT1-7)是一组NAD+依赖的去乙酰化以及ADP-核糖基转移酶家族,它们在能量代谢、基因组稳定性、衰老以及压力抵抗等方面起到重要作用[1]。SIRT7主要在细胞核表达,它能与组蛋白相互作用来调节核糖体基因转录[2]。 SIRT7被认为在转录过程中对连接染色体重建复合体与RNA Pol I中起到重要作用[3]。有研究发现SIRT7在小鼠模型和murine细胞系中有抑制肿瘤生长的作用[4]。但也有研究表明SIRT7能通过去乙酰化和抑制肿瘤抑制基因的转录来促进肿瘤的生长[5]。甲 状腺癌中,目前有相矛盾的结果,DE NIGRIS等[6]发现SIRT7在甲状腺癌细胞和组织中表达升高,不过ALJADA等[7]和CHEN等[8]却发现SIRT7在甲状腺癌中的mRNA表达水平低于癌旁非肿瘤性甲状腺组织,并且,到目前为止,SIRT7在甲状腺癌细胞中的作用未见报道。因此,本研究拟通过研究干扰SIRT7对甲状腺癌细胞BCPAP体外增殖、迁移和侵袭能力的影响,以进一步明确SIRT7与甲状腺癌的关系。
1 材料和方法
1.1 材料
1.1.1 载体和病毒:干扰SIRT7的慢病毒由上海汉恒生物科技有限公司生产。载体为pHBLV-CMVIEZsGreen-T2A-Puro。干扰慢病毒(shSIRT7组)和阴性对照病毒(阴性对照组)的最终病毒滴度均为2× 108PFU/mL。
1.1.2 细胞系:人甲状腺癌细胞系BCPAP购自中国科学院上海细胞生物学研究所。
1.1.3 主要试剂:10%胎牛血清、青霉素/链霉素、高糖DMEM培养基(美国Gibco公司);反转录试剂盒(Prime-ScriptTMRT Master Mix,日本TaKaRa公司);RIPA裂解液、BCA蛋白浓度试剂盒(上海碧云天公司);兔抗人SIRT7多克隆抗体(1:1 000,ABV11620-50,苏州百奇生物科技有限公司);山羊抗兔二抗(ab97200,英国Abcam公司);兔抗人GAPDH(1:1 000,ab8245,英国Abcam公司);辣根过氧化物酶HRP-ECL(美国Millipore公司);CCK-8试剂盒(日本Dojindo公司);Transwell板(美国Corning公司);侵袭胶(美国BD公司)。
1.2 方法
1.2.1 细胞培养:将细胞培养在含10%胎牛血清和青霉素/链霉素的高糖DMEM培养基中。CO2培养箱设定的条件为37 ℃,5% CO2。干扰SIRT7的稳定细胞株由慢病毒转染并在2 µg/mL的嘌呤霉素中筛选2周。
1.2.2 RT-PCR:用反转录试剂盒将500 ng总RNA合成cDNA。将cDNA稀释3倍后用RT-PCR试剂盒在RTPCR仪(美国BioRad公司)上合成。选用GAPDH为内参基因。各基因的引物为:SIRT7正向引物5’-AAAGGG AGAAGCGTTAGTGC-3’,反向引物5’-ACGCAGGAGGTACAG ACTTC-3’;GAPDH正向引物5’-TCAAGAAGGTGGTGAAGC AGG-3’,反向引物5’-TCAAAGGTGGAGGAGTGGGT-3’。PCR反应条件为:94 ℃ 2 min,然后94 ℃ 30 s,57 ℃ 30 s,72 ℃ 1 min,进行40个循环,最后72 ℃ 5 min后4 ℃维持。循环结束后,分析熔解曲线保证PCR产物的均一性。用2-ΔCt法对数据转换后进行统计分析。
1.2.3 Western blot:细胞用RIPA裂解液在冰上裂解30 min,然后离心收集上清并用BCA蛋白浓度试剂盒测定浓度后经聚丙烯酰胺凝胶电泳后转到PVDF膜上。将膜置于10%脱脂牛奶中室温封闭1 h后用一抗稀释液稀释的兔抗人SIRT7多克隆抗体在4 ℃冰箱孵育过夜。第2天用山羊抗兔二抗室温孵育30 min。二抗孵育结束后,用辣根过氧化物酶HRP-ECL发光法进行显色,在凝胶成像仪上拍摄[9]。内参抗体为兔抗人GAPDH。
1.2.4 细胞增殖活力检测:参照CCK-8试剂盒操作说明进行检测。实验前1 d将处于对数生长期的细胞以1 000个/孔的密度接种在96孔板。检测时向96孔板每孔中加入10 µL的CCK-8溶液,然后将培养板放入培养箱内孵育2 h后用酶标仪测定在450 nm 处测定吸光度。去掉每组5个复孔中的最大值和最小值,统计吸光度作为细胞增殖活力值。铺板第2天检测数值计为第1天。铺板后隔1天换1次培养基。细胞增殖活力(%)=[A(加药)-A(空白)]/ [A(未加药)-A(空白)]×100%。A(加药):具有细胞、CCK-8溶液和药物溶液的孔的吸光度;A(空白):具有培养基和CCK-8溶液而没有细胞的孔的吸光度;A(未加药):具有细胞、CCK-8溶液而没有药物溶液的孔的吸光度。
1.2.5 划痕实验:干扰SIRT7的BCPAP细胞和阴性对照细胞以5×105每孔的密度接种在6孔板中并培养过夜。当细胞密度接近90%时,用移液器枪头在孔底的细胞层上划痕,然后用PBS缓冲液冲洗细胞3遍以移除脱落的细胞,剩余细胞在无血清培养基中培养,继续培养24 h后在显微镜下拍照并测量细胞迁移距离。
1.2.6 细胞迁移和侵袭实验:迁移实验:将6×104个细胞混悬在0.2 mL无血清培养基中并铺板在24孔Transwell板的上层小室中,下层小室加入0.6 mL 含10%胎牛血清的培养基。细胞在37 ℃培养箱培养18 h,然后用棉签将小室上层的细胞擦除,将小室底部的细胞用甲醛固定后再用0.1%结晶紫染液染色,在显微镜下对小室底部的迁移细胞进行观察拍照。侵袭实验与迁移实验步骤相似,但需要在细胞铺板前在上层小室中加入侵袭胶。
1.3 统计学处理方法 采用SPSS20.0统计软件进行统计分析。计量资料以±s表示,2组间比较采用独立样本t检验。P<0.05为差异有统计学意 义。
2 结果
2.1 构建干扰SIRT7的甲状腺癌细胞BCPAP稳定株 分别在mRNA和蛋白水平验证SIRT7的干扰结果。shSIRT7组的mRNA水平较阴性对照组显著下降(P<0.01),shSIRT7组细胞的SIRT7蛋白表达量也较阴性对照组显著降低(P<0.01),见图1。
2.2 干扰SIRT7对BCPAP细胞系的增殖、迁移和侵袭能力的影响 干扰SIRT7之后,BCPAP细胞的增殖能力显著降低,划痕愈合能力也下降,迁移和侵袭能力都受到了显著抑制,差异均有统计学意义(P<0.05),见图2-4。

图1 干扰SIRT7后BCPAP细胞的SIRT7 mRNA和蛋白相对表达量

图2 干扰SIRT7后BCPAP细胞的体外增殖活力变化
3 讨论
研究显示,多个SIRT家族成员在不同的肿瘤中扮演了不同的角色,这可能取决于具体的组织和肿瘤类型[10]。比如SIRT1在胃癌[11]、结肠癌[12]、前列腺癌[13]以及皮肤癌[14]等肿瘤中表达是升高的,这提示它在这些肿瘤中应该扮演了促进肿瘤形成的作用。但另外有一些研究发现SIRT1在乳腺癌[15]中表达是降低的,并且,在小鼠APCmin/+模型中,SIRT1能抑制小鼠肠道肿瘤的形成[16]。与此相类似的还有SIRT2,它在乳腺癌[17]、胶质瘤[18]、皮肤癌[19]等肿瘤中表达降低,而在急性髓性白血病[20]及前列腺癌[21]中表达升高。因此,我们不能随便把一种肿瘤中的研究结论外推到另一种肿瘤中去。

图3 干扰SIRT7后BCPAP细胞的划痕愈合能力变化
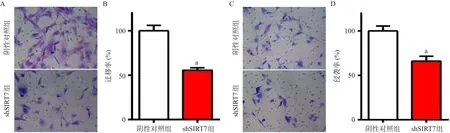
图4 干扰SIRT7后BCPAP细胞的迁移和侵袭能力变化
目前发现SIRT7在大多数肿瘤中的表达是上调的。比如KIM等[22]发现SIRT7在人肝癌组织中表达上调并且敲减SIRT7能通过影响细胞周期和自噬相关蛋白来抑制肝癌细胞体外和体内生长。YU等[23]发现SIRT7在结肠癌中表达上调并且SIRT7能通过调节MAPK信号通路和EMT来影响结肠癌细胞的增殖和迁移。ASHRAF等[24]发现SIRT7在乳腺癌中表达上调,GENG等[25]发现SIRT7表达上调与乳腺癌的不良预后有关。此外,SIRT7还被发现在胃癌[26]和卵巢癌[27]细胞中表达上调。MCGLYNN等[28]等却发现SIRT7在胰腺癌中表达降低,并且SIRT7表达高的胰腺癌患者有更长的术后生存时间。这些结果提示SIRT7可能同时具有癌基因和抑癌基因的作用。目前SIRT7在甲状腺癌中的研究有不一致的结论,DE NIGRIS等[6]发现SIRT7在甲状腺癌细胞和甲状腺癌组织中表达升高,但是ALJADA等[7]在包含乳腺癌、结肠癌、肝癌、甲状腺癌等多个肿瘤标本的cDNA芯片上研究了SIRT7的表达情况,发现SIRT7的mRNA表达水平在甲状腺癌中是显著下调的。我们之前的研究通过对包含43例甲状腺癌和癌旁甲状腺组织的组织芯片进行免疫组织化学实验,发现SIRT7在甲状腺癌中的蛋白水平低于癌旁非肿瘤甲状腺组织[8]。
本研究发现干扰SIRT7能显著抑制BCPCP细胞的增殖、迁移和侵袭能力。这些结果表明SIRT7可能在甲状腺癌中扮演了癌基因的角色,因此,SIRT7可能是甲状腺癌一个潜在的治疗靶点。
猜你喜欢
承德医学院学报(2022年2期)2022-05-23
东坡赤壁诗词(2021年2期)2021-06-01
国际放射医学核医学杂志(2021年10期)2021-02-28
国际放射医学核医学杂志(2021年10期)2021-02-28
中华养生保健(2020年4期)2020-11-16
奥秘(创新大赛)(2019年9期)2019-10-09
环球时报(2019-01-29)2019-01-29
畅谈(2018年6期)2018-08-28
奥秘(2017年5期)2017-07-05
癌变·畸变·突变(2015年3期)2015-02-27
