
洛阳烟区烟株根围抗烟草疫霉放线菌的筛选与鉴定
2019-01-25 03:18陈奇园丁钥琪赵世民李淑君康业斌
烟草科技 2019年1期
陈奇园,丁钥琪,赵世民,李淑君,康业斌*
1.河南科技大学林学院,河南省洛阳市开元大道263号 471023
2.河南省烟草公司洛阳市公司,河南省洛阳市开元大道246号 471023
3.河南省农科院许昌烟草研究所,河南省许昌市魏都区永昌大道 461000
由烟草疫霉(Phytophthora parasitica var.nicotianae)侵染烟草引起的黑胫病是一种分布广泛、危害严重的毁灭性土传真菌病害[1]。目前,烟草黑胫病的防治措施主要采用种植抗病品种、合理轮作与药剂防治。然而,随着烟草栽培制度的改变,抗病品种连年、大面积单一种植,极易导致其抗病性丧失。生产上常用的甲霜灵等几种内吸性杀菌剂对烟草疫霉作用位点单一,容易产生抗药性而引起药效下降,从而导致药剂用量加大,使烟叶中的农药残留增加、造成环境污染[2-4]。近年来,国内外学者相继开展了拮抗放线菌防治烟草病害的研究。Lukic等[5]从烟田土壤中分离出28株放线菌,其中两株对12种烟草病原真菌有拮抗作用;Kortemaa等[6]报道了世界上已广泛应用的一种放线菌的活体制剂防治腐霉菌、疫霉菌、镰刀菌和丝核菌等常见的土传病原菌;Loqman等[7]从摩洛哥葡萄根际土壤中分离获得142株放线菌,并测定了对尖孢镰刀菌、终极腐霉、大丽轮枝菌、灰葡萄孢菌及白绢病菌的拮抗作用,其中有24株放线菌至少抗4种病原菌。周喜新等[8]从湖南长沙浏阳市北盛镇烟草黑胫病发病田健株根际土壤中分离纯化获得1株对烟草疫霉具有显著拮抗作用的放线菌LY18,其发酵液对烟草疫霉菌丝的抑制率达70%;王静等[9]从山东省日照市五莲县于里镇烟田健株根际土壤分离纯化获得1株拮抗放线菌F8,室内测定对供试烟草疫霉具有较强的抑制作用,温室盆栽试验防效为70.3%。但河南省烟田土壤中拮抗放线菌种类及开发应用尚未见系统研究报道,为此,进行了洛阳地区烟草根围土壤中拮抗放线菌的筛选与鉴定,旨在为进一步开发利用拮抗微生物提供依据。
1 材料与方法
1.1 试验材料
烟 草 疫 霉(Phytophthoraparasiticavar.nicotianae)以及从烟草根围土壤分离的放线菌稀释涂布平板均由河南科技大学植物病害分子鉴定与绿色防控实验室保存。
试验所用培养基包括:高氏一号琼脂培养基、马铃薯葡萄糖琼脂培养基、大豆酵母培养基、淀粉铵琼脂培养基、蔗糖察氏琼脂培养基、葡萄糖天门冬素琼脂培养基、克氏一号琼脂培养基和葡萄糖酵母膏琼脂培养基。细菌基因组DNA快速抽提试剂盒(编号:B518255)和 PCR扩增试剂盒(编号:B532491)购自生工生物工程(上海)股份有限公司。
1.2 试验方法
1.2.1 根围放线菌的纯化
将高氏一号培养基融化,待冷却至50℃左右,加入3%的重铬酸钾溶液使培养基中重铬酸钾浓度约100 mg/L,倒平板;待平板凝固后备用,用灭菌牙签从前期试验保存的稀释涂布平板上挑出菌落形态或颜色不同的放线菌单菌落至高氏一号平板上划线分离、纯化培养[10],直至获得单菌落。
1.2.2 拮抗菌株的筛选
1.2.2.1 拮抗菌株的初筛
采用改良的平板对峙培养法[11]测定菌株对烟草疫霉的拮抗作用,保留抑菌率≥50%的菌株进行复选。
1.2.2.2 拮抗菌株的复筛
采用菌丝生长速率法[12]测定初选菌株发酵液对烟草疫霉的抑菌活性,选取抑菌率达100%的菌株进一步鉴定其种类。
1.2.3 拮抗菌株的鉴定
1.2.3.1 形态及培养特征观察
用插片法[13]在普通光学显微镜(无锡瀚光光学科技有限公司)100倍油镜下观察记载其基内菌丝体有无横隔、是否断裂;气生菌丝体的特征;孢子链的形状、着生方式;孢子的形状等。
根据《链霉菌鉴定手册》[14],将菌株分别接种在高氏一号琼脂培养基、马铃薯葡萄糖琼脂培养基、淀粉铵琼脂培养基、蔗糖察氏琼脂培养基、葡萄糖天门冬素琼脂培养基、克氏一号琼脂培养基、葡萄糖酵母膏琼脂培养基上,28℃下培养,分别在7、14、28 d后观察菌株在不同培养基上气生菌丝、基内菌丝的生长状况,是否有可溶性色素产生及其颜色。
1.2.3.2 生理生化特征观察
参照《植病研究法》[12]和《放线菌的分类和鉴定》[15]进行明胶液化、牛奶凝固和胨化、淀粉分解、纤维素水解、硫化氢与黑色素产生以及利用碳源产酸等试验,观察生理生化特征。
1.2.3.3 16S rDNA序列分析
按照细菌基因组DNA快速抽提试剂盒(编号:B518255)说明书进行放线菌DNA的提取。以提取的DNA为模板,采用通用引物为27F:5′-AGAGTTTGATCCTGGCTCAG-3′和1492R:5′-GGTTACCTTGTTACGACTT-3′对放线菌16S rDNA序列进行PCR扩增。PCR反应体系(50 μL):模板DNA 1.0 μL,10×Taq Buffer(NH4+)5.0 μL,dNTP Mix(10.0 mmol/L)1.0 μL ,Taq DNA Polymerase(5 U/μL)0.25 μL,MgCl2(25 mmol/L)3.0 μL,上下游引物(10.0 μmol/L)各 1.0 μL,加 dd H2O 补至 50 μL。PCR反应条件:94℃预变性2 min;94℃变性1 min,55 ℃退火30 s,72 ℃延伸2 min,30个循环,最后72℃延伸10 min,4℃保存。检测到合适的条带后,将其所对应的原始扩增产物(未纯化,45 μL)由生工生物工程(上海)股份有限公司测序。将测序得到的序列在NCBI网站上进行BLAST比对,用MEGA.6软件的邻接法构建系统发育树[16]。
2 结果与分析
2.1 根围放线菌的纯化
共挑出放线菌单菌落496个,进一步分离纯化获得形态或颜色不同的放线菌单菌落227个。
2.2 拮抗菌株的筛选
2.2.1 拮抗菌株的初筛
表1显示,有45株放线菌对烟草疫霉的抑菌率大于50%,其中25株的抑菌率大于70%。

表1 放线菌对烟草疫霉的抑制作用①Tab.1 Inhibition effects of actinomycetes against P.parasitica
2.2.2 拮抗菌株的复筛
表2表明,15株拮抗放线菌的发酵液对烟草疫霉的抑菌率大于50%。其中,菌株YY75、YY124、LN204、LN221、LN247、SX92、YY135 和SX59的抑菌率达100%,确定为优势拮抗菌株。
2.3 优势拮抗菌株的鉴定
2.3.1 形态及培养特征观察
菌株YY75孢子丝弯曲,孢子圆形;菌株LN204、LN221和LN247孢子丝螺旋形,孢子圆柱状;菌株SX92孢子丝波曲,孢子杆状;菌株YY124和YY135孢子丝圈卷,孢子杆状;菌株SX59孢子丝螺旋形,孢子杆状(图1)。
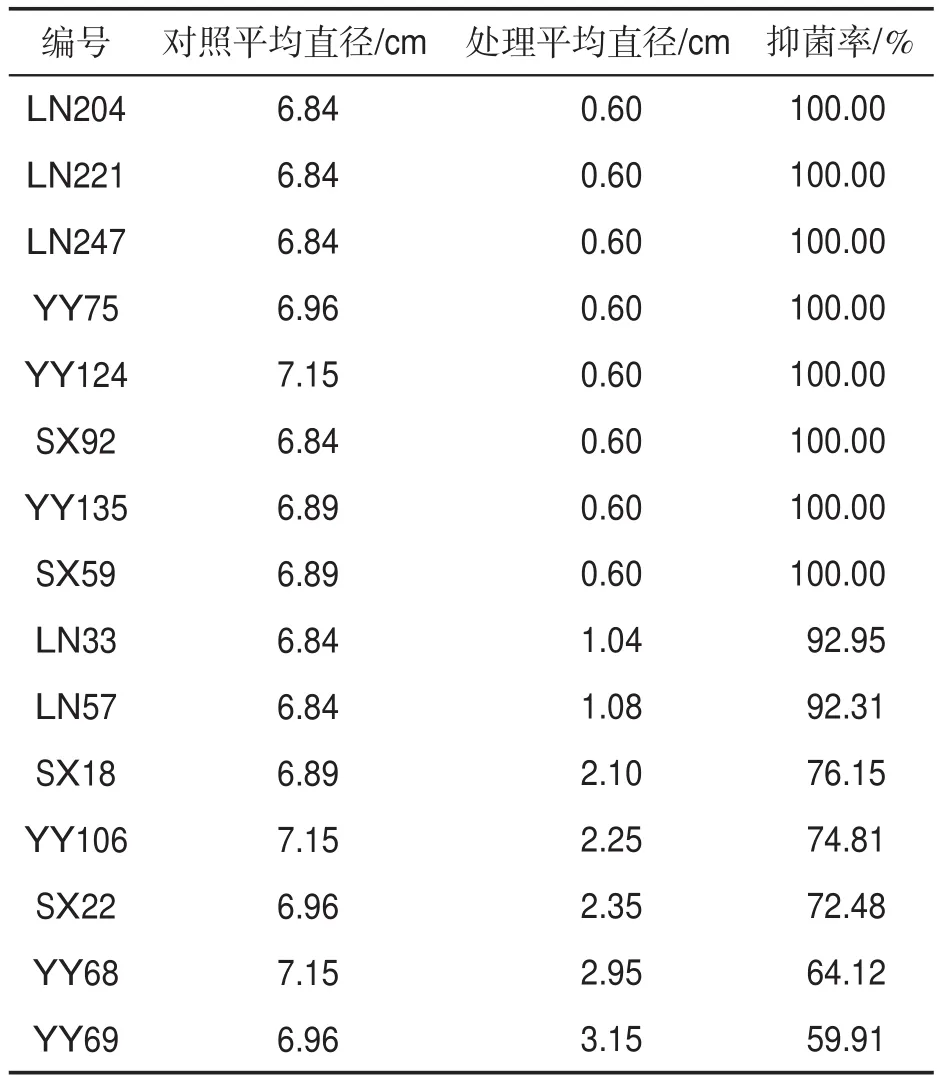
表2 拮抗放线菌对烟草疫霉的抑制作用Tab.2 Inhibition effects of antagonistic actinomycetes against P.parasitica
在高氏一号琼脂培养基上,菌株YY75气生菌丝白色,基内菌丝乳脂色至紫红色,无可溶性色素;菌株LN204、LN221和LN204气生菌丝白至灰色,基内菌丝蜜黄色,无可溶性色素;菌株SX92气生菌丝白色,基内菌丝乳脂色,无可溶性色素;菌株YY124和YY135气生菌丝白色至粉红色,基内菌丝紫红色,无可溶性色素;菌株SX59气生菌丝白色至粉红色,基内菌丝紫红色,无可溶性色素。
在马铃薯葡萄糖琼脂培养基上,菌株YY75气生菌丝白色,基内菌丝乳脂色至褐色,可溶性色素黄色;菌株LN204、LN221和LN204少量灰色气生菌丝,基内菌丝米黄色,无可溶性色素;菌株SX92气生菌丝白色,基内菌丝褐绿至橄榄灰绿色,可溶性色素墨绿色;菌株YY124和YY135无气生菌丝,基内菌丝紫红色,无可溶性色素;菌株SX59气生菌丝粉红色,基内菌丝紫红色,可溶性色素米色。
在淀粉铵琼脂培养基上,菌株YY75无气生菌丝,基内菌丝米绿色,无可溶性色素;菌株LN204、LN221和LN204气生菌丝白至鼠灰色,基内菌丝微褐,无可溶性色素;菌株SX92无气生菌丝,基内菌丝米绿色,无可溶性色素;菌株YY124和YY135气生菌丝粉红至石南紫,基内菌丝玫瑰色至紫红色,无可溶性色素;SX59气生菌丝古粉红色,基内菌丝玫瑰色至信号红,无可溶性色素。
在蔗糖察氏琼脂培养基上,菌株YY75无气生菌丝,基内菌丝乳脂色至米绿色,无可溶性色素;菌株LN204、LN221和LN204气生菌丝沙紫灰至暗灰色,基内菌丝浅灰至灰黄色,无可溶性色素;菌株SX92气生菌丝白色,基内菌丝硫磺色,可溶性色素黄色;菌株YY124和YY135气生菌丝白色至粉红色,基内菌丝橘红至宝石红,无可溶性色素;菌株SX59气生菌丝白色至粉红色,基内菌丝橘红至宝石红,无可溶性色素。
在葡萄糖天门冬素琼脂培养基上,菌株YY75无气生菌丝,基内菌丝乳脂色至紫红色,无可溶性色素;菌株LN204、LN221和LN204无气生菌丝,基内菌丝无色至微黄,无可溶性色素;菌株SX92无气生菌丝,基内菌丝乳脂色,无可溶性色素;菌株YY124和YY135气生菌丝白色,基内菌丝石南紫至酒红紫,无可溶性色素;菌株SX59气生菌丝白色,基内菌丝酒红紫,无可溶性色素。
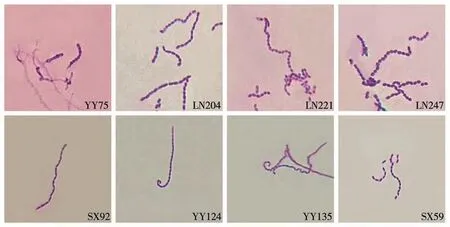
图1 菌株的孢子丝和孢子形态(×100)Fig.1 Spore chains and spore morphology of antagonistic strains(×100)
在克氏一号琼脂培养基上,菌株YY75气生菌丝白色,基内菌丝乳脂色至葡萄酒红,无可溶性色素;菌株LN204、LN221和LN204气生菌丝浅黄色至灰色,基内菌丝蜜黄色,无可溶性色素;菌株SX92气生菌丝白色,基内菌丝乳脂色,无可溶性色素;菌株YY124和YY135气生菌丝粉红至古粉红色,基内菌丝粉红至紫红色,无可溶性色素;菌株SX59气生菌丝白色至粉红色,基内菌丝粉红至橘红,无可溶性色素。
在葡萄糖酵母膏琼脂培养基上,菌株YY75无气生菌丝,基内菌丝为象牙色至赭黄色,可溶性色素为黄棕色;菌株LN204、LN221和LN204气生菌丝为白至灰色,基内菌丝米为褐色,无可溶性色素;菌株SX92气生菌丝为白色,基内菌丝为米褐色,可溶性色素为棕色;菌株YY124和YY135气生菌丝为白色,基内菌丝为象牙色至火焰红,无可溶性色素;菌株SX59气生菌丝为粉红色,基内菌丝也为粉红色,无可溶性色素。
2.3.2 生理生化特性
优势拮抗菌株均可利用葡萄糖、麦芽糖、阿拉伯糖和甘露醇等多种碳源,明胶液化、纤维素水解均为阳性,而硫化氢的产生均为阴性;菌株YY135对牛奶的凝固和胨化反应为阳性;菌株YY75和SX92黑色素的产生反应也为阳性,见表3。
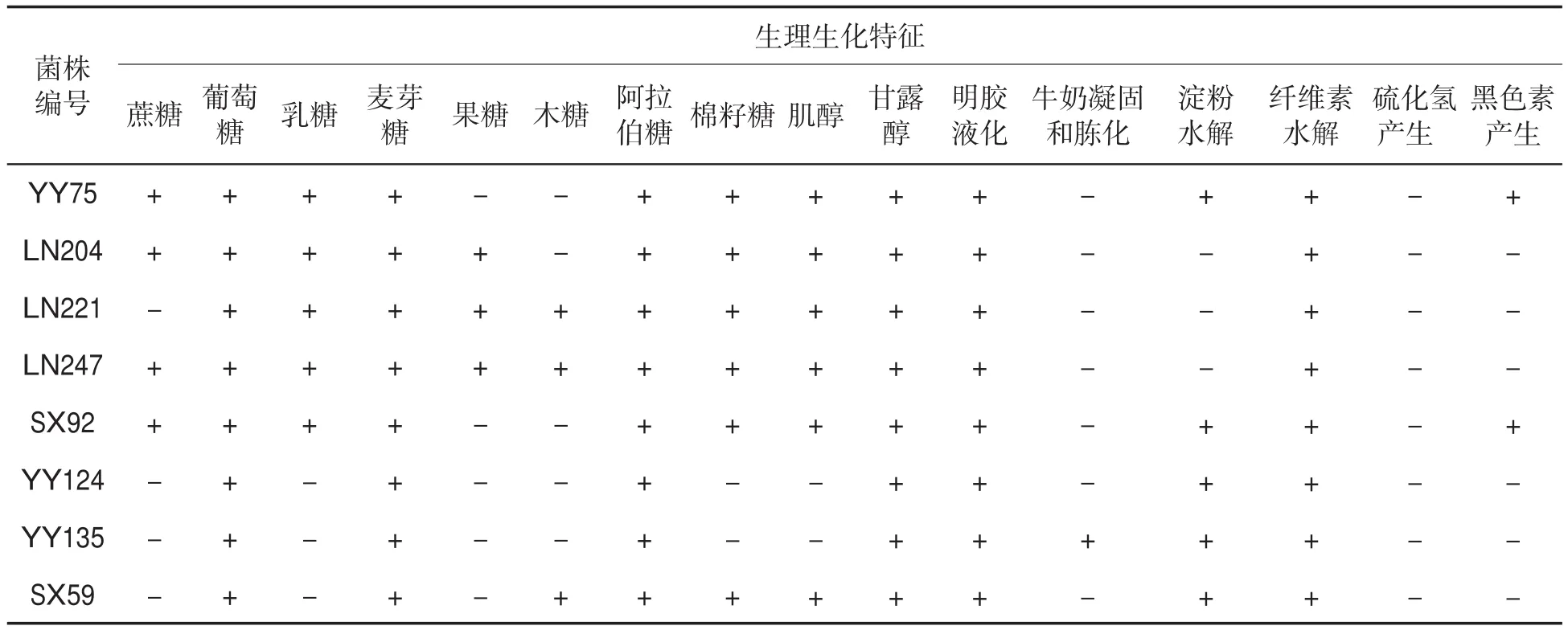
表3 优势拮抗菌株的生理生化特征①Tab.3 Physiological and biochemical characteristics of dominant antagonistic strains
2.3.3 16S rDNA序列分析
将菌株LN204、LN221、LN247、SX59、YY124、YY135、YY75和SX92的16S rDNA序列提交至NCBI,登 录 号 分 别 为 MH265958、MH265959、MH265960、MH265965、MH265966、MH265967、MH265968和MH265970。进一步对8个菌株的16S rDNA序列进行BLAST比对,用MEGA6软件构建系统发育树(图2)。结果表明,菌株YY75的序列与登录号为NR043827(Streptomyces lucensis)等序列同源性大于99%,菌株YY124和YY135的序列与登录号为AB184370(S.nobilis)等序列同源性大于99%,菌株LN204、LN221和LN247的序列与登录号为JX284411(S.rochei)等序列同源性大于99%,菌株SX92的序列与登录号为AB184197(S.coralus)等序列同源性大于99%,菌株SX59的序列与登录号为 AB249919(S.aureoverticillatus)等序列同源性大于99%。
3 讨论
本试验中鉴定的8株优势拮抗放线菌均属于链霉菌属,这与边传红[17]从洛阳烟草根际土壤中筛选鉴定的13株拮抗放线菌也均属于链霉菌属的结果一致;也与廖振林等[18]从广西北海红树林土壤中筛选鉴定的10株拮抗放线菌,其中有8株属于链霉菌属的结果一致。说明链霉菌属是土壤放线菌中常见的属。本试验中获得的优势拮抗放线菌发酵液对烟草疫霉抑制率均达100%,说明发酵液中含有抑菌活性成分,这与Singh等[19]发现的S.lucensis可以产生鲁萨霉素C和Anukool等[20]发现的S.rochei F20可以产生抗生素Streptothricin的试验结果一致,说明链霉菌属是放线菌中主要产抗生素的属。但有关链霉菌属发酵液中活性成分分析、抑菌机理的研究以及盆栽防效试验与田间防治效果等仍有待进一步的深入研究。
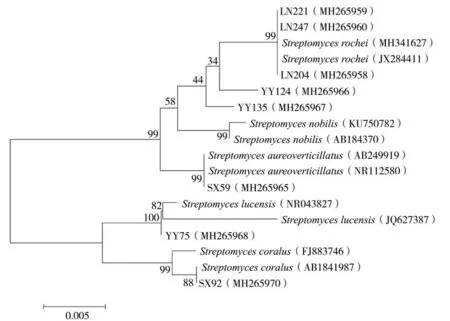
图2 优势拮抗菌株16S rDNA基因序列构建的系统发育树Fig.2 Phylogenetic tree of dominant antagonistic strains based on 16S rDNA sequences
4 结论
从洛阳烟区烟株根围土壤中分离纯化共获得227个放线菌单菌落,其中8株的发酵液对烟草疫霉菌丝生长的抑制率达100%,被确定为优势拮抗放线菌。根据这些菌株的形态学观察、生理生化反应和16S rDNA序列分析结果,菌株YY75鉴定为鲁萨链霉菌(S.lucensis),菌株YY124和YY135鉴定为高贵链霉菌(S.nobilis),菌株LN204、LN221和LN247鉴定为娄彻链霉菌(S.rochei),菌株SX92鉴定为珊瑚链霉菌(S.coralus),菌株SX59鉴定为金黄回旋链霉(S.aureoverticillatus),均属于放线菌门、放线菌纲、链霉菌目、链霉菌科、链霉菌属。
猜你喜欢
食品与发酵工业(2022年18期)2022-10-04
陕西农业科学(2022年2期)2022-03-28
新农业(2021年9期)2021-06-20
浙江农业学报(2021年5期)2021-05-31
食品工业科技(2019年21期)2019-11-27
中小学实验与装备(2019年3期)2019-07-03
天然产物研究与开发(2018年10期)2018-11-06
新农业(2017年15期)2017-09-11
上海农业学报(2017年4期)2017-04-10
天然产物研究与开发(2014年7期)2014-04-27
