
2017年南京市2株肠道病毒71型分离株全基因组序列测定分析
2019-01-21 02:05:50,,,
中国人兽共患病学报 2018年12期
, ,,
手足口病(hand,foot and mouth disease,HFMD)是一种儿童常见的自限性传染病,多发于1~5岁的儿童,其临床症状为婴幼儿发热和手、足、口腔和臀等处的皮疹或疱疹[1-2]。引起HFMD的病原体有多种,其中以肠道病毒71型(Enterovirus 71,EV71)和柯萨奇病毒A16型(Coxsackievirus A16,CA16)最为常见[3]。EV71属于小RNA病毒科、肠道病毒属A组的成员,为单股正链RNA病毒。基因组全长约7 400 bp,位于基因组两端的5′和3′非编码区(untranslated regions,UTRs)在病毒的复制和蛋白的翻译中发挥着重要作用[4]。病毒基因组仅含有一个开放阅读框(open reading frame,ORF),编码一个由2 193个氨基酸组成的多聚蛋白,分为结构前体蛋白P1和非结构前体蛋白P2、P3。P1蛋白可进一步加工为结构蛋白VP1~VP4,其中VP1区是病毒基因分型的主要依据。一般情况下,儿童感染EV71与感染CA16病毒后的临床症状不易区分[5],但相对而言,EV71感染所引起的HFMD可能伴有神经性肺水肿和急性弛缓性麻痹、无菌性脑膜炎等神经系统并发症,严重地危害了婴幼儿的健康[1, 6]。
近年来EV71在国内的流行呈显著上升趋势,其中以C4基因型为主[7]。在江苏省境内每年都能检测出EV71病毒的疫情,比对这些毒株的序列后发现,流行的毒株主要属于C4a亚型[8]。因此加强对中国大陆各地区EV71流行毒株的基因特征分析,对于有效地预防和控制手足口病的流行具有重要的意义。本文通过从患者粪便样品中分离出的2株EV71病毒,对其进行全基因组序列比对、基因分型和种系进化关系分析等研究,以期为EV71的变异研究提供依据。
1 材料与方法
1.1 材料
1.1.1标本来源 EV71感染粪便标本收集自南京市儿童医院2017年5-7月经临床诊断为EV71感染手足口病的入院治疗患儿。粪便采集后立即分装至2 mL冻存管,于-80 ℃低温冷冻保存。
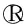
1.2 方法
1.2.1粪便中EV71的分离、复制 取粪便标本0.3 g,加入1.5 mL PBS制成20%粪便悬液,充分混匀,12 000 r/min离心20 min。取上清液1 mL至新的离心管,加入100 μL氯仿,充分混匀,12 000 r/min离心15 min,离心后取200 μL上清加入2.8 mL含有2%FBS DMEM中,接种于致密单层的Vero细胞上。细胞置于37 ℃、5%CO2的细胞培养箱中培养,接种后每天观察细胞病变效应(CPE),若出现CPE,则取100 μL阳性上清液传至新的Vero细胞中,2 h后吸弃上清液,换成含有2%FBS DMEM,继续观察细胞CPE变化。CPE达到75%以上时,收集病变细胞液,-80 ℃保存。如7 d后没有出现CPE,盲传3代,仍未见CPE出现,则判为阴性。
1.2.2引物设计与合成 引物设计参考文献[9]。选取GenBank中EV71各地区代表株的全基因组序列,经序列比对后找出病毒基因序列的保守区域,采取分段扩增方式依序设计8对首尾相互重叠、覆盖基因组全长的引物(见表1)。引物序列由通用生物系统有限公司合成。

表1 EV71全基因组分段扩增和测序的引物Tab.1 Primers used for fractional amplification and sequencing of whole genome of EV71
1.2.4序列测定与数据分析 8段特异性扩增得到的PCR产物送至通用生物系统有限公司进行序列测定。通过Chromas软件显示各段产物依序位置的碱基峰图对测序所得的序列进行可靠性分析,通过SeqMan软件将比对的序列依序组装成一条完整的全基因序列。同时从NCBI基因数据库(GenBank)中下载EV71各基因型代表毒株序列共24条,见表2。应用Megalign软件对EV71分离毒株的核苷酸和氨基酸序列进行同源性比对分析,应用MEGA 7.0软件构建系统进化树(Bootstrap值为1 000)。
表2 序列分析中所用EV71毒株信息Tab.2 Background of EV71 strains in sequence analysis
2 结 果
2.1病毒分离 将临床检测为EV71-RNA阳性的36份手足口病患者的粪便标本,处理后分别接种至致密生长的Vero细胞,培养分离出2个阳性分离物。阳性分离物再次接种Vero细胞,培养2 d后,镜下观察可见细胞圆缩、透亮、聚堆的病变现象,见图1。
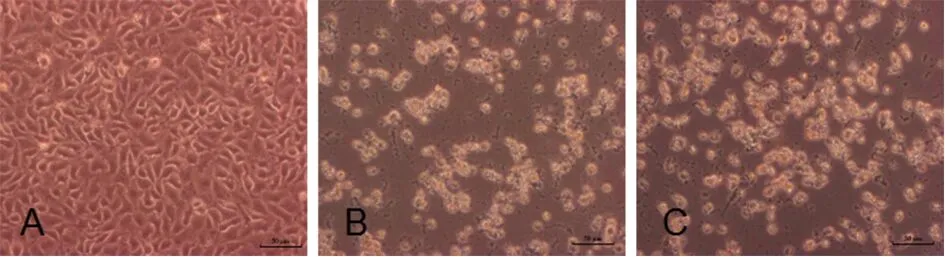
A: Control Vero cells;B: CPE on Vero cells infected with NJ2017iso1 after 48 h;C: CPE on Vero cells infected with NJ2017iso2 after 48 h图1 Vero细胞感染NJ2017分离株后的细胞病变(×200)Fig.1 CPE on Vero cells infected with EV71 isolates
2.2PCR产物鉴定 EV71阳性粪便分离物感染Vero细胞后提取RNA,采用8对特异性设计的引物进行RT-PCR扩增,PCR产物经1.5%浓度琼脂糖凝胶电泳,显示各条扩增片段均与引物设计的理论扩增长度相符,见图2。
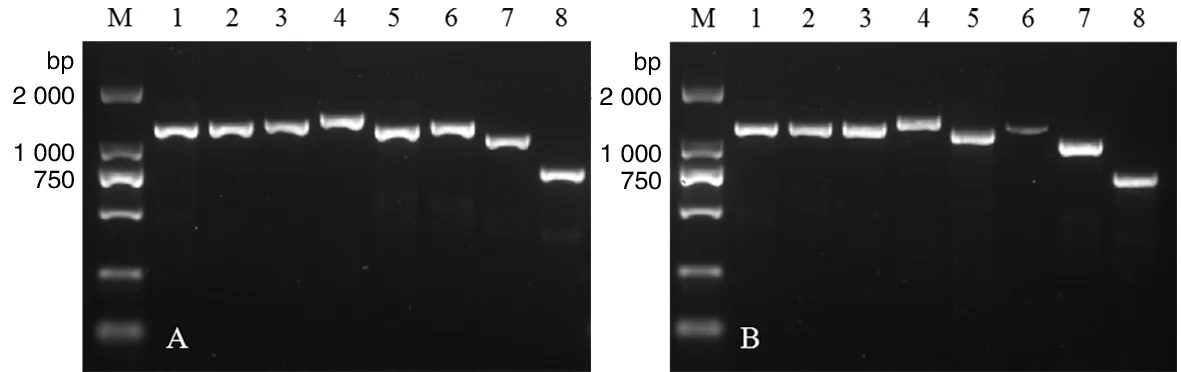
M: DNA Marker DL2000; A: EV71-NJ2017iso1 infected-products were amplified fractionally by PCR with 8 pairs of specific primer; B: EV71-NJ2017iso2 infected-products were amplified fractionally by PCR with 8 pairs of specific primer图2 NJ2017分离株8对引物PCR产物电泳图Fig.2 Electropherogram of PCR products of EV71 isolates
2.3基因组序列的测定及分析 经分段扩增,序列测定和比对拼接后,获得NJ2017 iso1和NJ2017 iso2的全基因组序列,并将序列提交至GenBank数据库,基因序列号分别为MG581490和MG520666。EV71-NJ2017 iso1基因组全长为7 365 bp,其中5’UTR区长为692 bp,3′UTR区为89 bp;NJ2017 iso2基因组全长为7 358 bp,其中5′UTR长为705 bp,3′UTR区长81 bp。两株EV71基因组编码区均为6 582个核苷酸,仅含有一个开放阅读框,编码一含2 193个氨基酸残基的多聚蛋白(polyprotein),经基因组序列比对,两分离株核苷酸同源性达到94.0%。从表3可看出,NJ2017 iso2与EV71 BrCr原型株基因组的核苷酸和氨基酸同源性分别为79.8%和95.0%。
2.4基因组核苷酸序列同源性比对 将NJ2017 iso1,2株与来自GenBank中不同EV71代表株的核苷酸序列进行同源性分析,选择NJ2017 iso2作为参照。从表3中可以看出,无论是全基因组,或是在5′UTR,衣壳蛋白(VP1,VP4),非衣壳蛋白(2A,3C,3D)的各个分区中,NJ2017 iso2与Fuyang 17.08-02的核苷酸同源性最高,为96.9%,与EV71 BrCr的同源性最低,为79.8%。同时还发现,南京市分离株之间的5′UTR和3D区存在显著的差异性,分别为74.4%和37.6%。
2.5编码蛋白氨基酸序列同源性比较 NJ2017 iso1,2株病毒基因组编码区全长6 582个核苷酸,共编码2 193个氨基酸。将NJ2017 iso2株与其他EV71型毒株氨基酸序列进行同源性比对,见表3。从编码的氨基酸组成成分分析,NJ2017 iso2株与Fuyang 17.08-02的氨基酸同源性最高,与NJ2017 iso1株同源性最低。虽然在核苷酸序列上,NJ2017分离株与这24株病毒株序列之间的差异变化显著,但氨基酸序列的同源性均高达95%以上,说明EV71病毒基因组中大多数核苷酸序列变异属无义突变,相应氨基酸序列的遗传变化相对稳定。
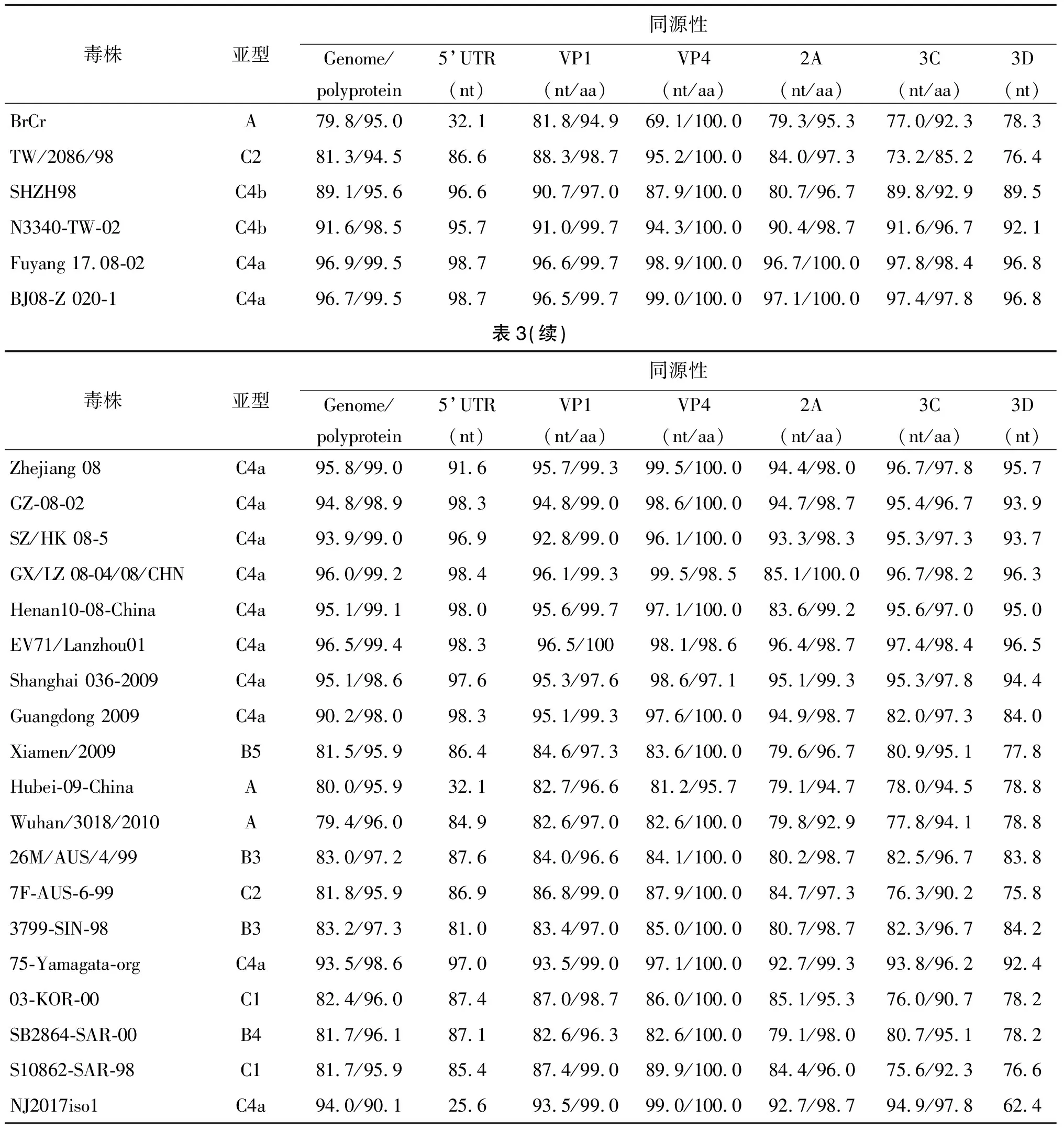
表3 南京市分离株NJ2017 iso2与其它分离株的核苷酸(nt)及氨基酸(aa)的同源性比较(%)Tab.3 Homology comparison of the nucleotide and amino acid sequence between NJ2017 iso2 and other EV71 isolates(%)
2.6病毒基因的种系进化分析 从GenBank上下载24株中国大陆各省市和亚太国家及地区的EV71代表株的全基因组序列,使用MEGA 7.0软件通过邻位归并法(Neighbor-joining),基于病毒外壳蛋白VP1基因和全基因组核苷酸序列,对NJ2017 iso1,2株与25株代表株进行遗传进化分析。无论在VP1基因(图3)还是在全基因组序列(图4)水平上构建的进化树中均可以看出,NJ2017 iso1,2株与C4亚型Fuyang 17.08-2、BJ08-Z020-1、Lanzhou01等株进化关系密切,同属于C4a进化分支,而与A型BrCr国际标准株、B亚型Xiamen/2009、26M/AUS/4/99、3799-SIN-98等株在进化上较远,这与核昔酸序列同源性比对的结果相符。NJ2017 iso1,2株与中国大陆各省市分离的10株代表株之间进化关系较近,分析结果与有关国内EV71主要代表毒株的基因型是C4亚型相符[10]。

图3 NJ2017 iso1,2株VP1序列遗传进化树Fig.3 Phylogenetic tree based on the VP1 gene between NJ2017 iso1,2 and the representatives of each subgenotype
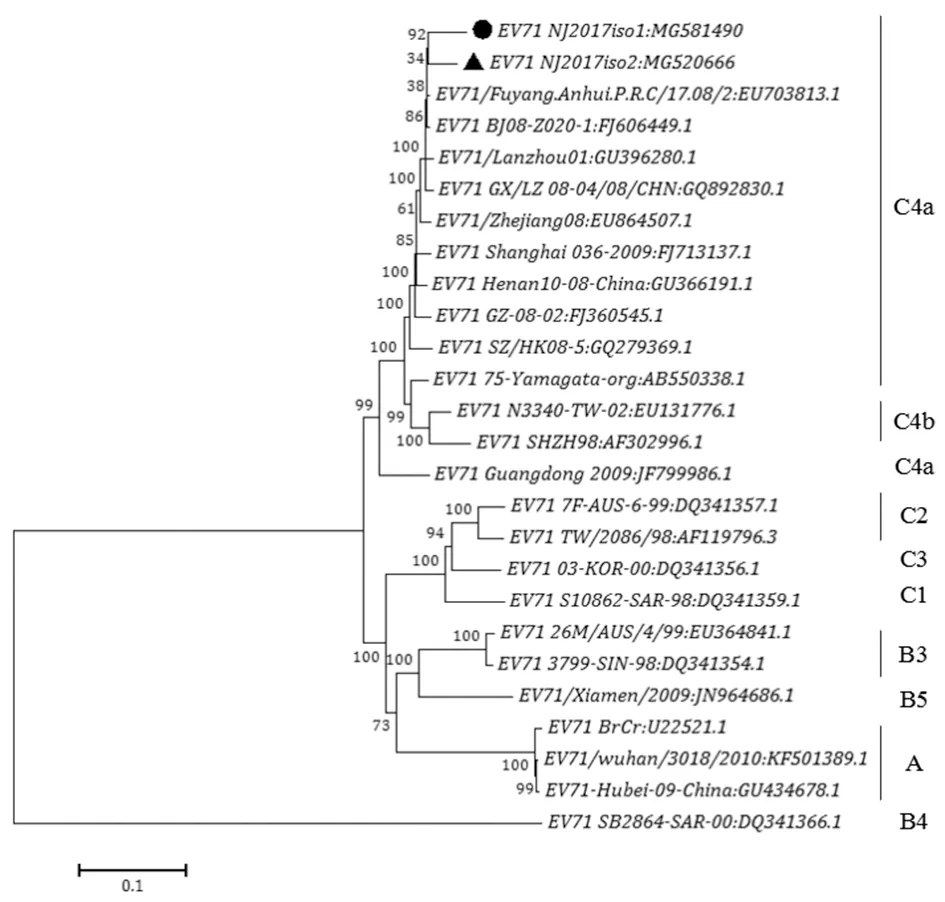
The representative strains and its Genbank accession numbers are listed in Phylogenetic tree,which are shown on the branches as Bootstrap values图4 NJ2017iso 1,2株全基因组序列遗传进化树Fig.4 Phylogenetic tree based on the full-length genome between NJ2017iso 1,2 and the representatives of each subgenotype
3 讨 论
近年来,EV71已逐渐成为继脊髓灰质炎病毒后又一重要的嗜神经肠道病毒。自1969年首次在美国加州被报道以来,EV71病毒已经在世界范围内引起多次大规模的暴发,但不同国家和地区的流行毒株的基因型存在显著差异。如1997-1999年新加坡、澳大利亚等亚太国家是以B3亚型感染为主;台湾省在1998年以C2亚型感染为主,而2000年之后逐渐被B4亚型取代。2008年以来在中国大陆的EV71感染引发的HFMD流行中,神经系统并发症的发病率逐年上升,因而加强对EV71毒株基因型的特征分析和流行病学的监控,在HFMD的预防及控制中有着重要的临床价值。本文通过对南京市2017年分离的2株EV71型病毒株的全基因组进行了序列测定、同源性分析及亲缘进化关系等方面的研究,为了解目前南京地区流行的EV71毒株的病原特征及遗传变异情况提供了重要的信息。
EV71病毒基因组仅含有1个开放阅读框,编码的一个多聚蛋白在自身蛋白酶的酶解作用下形成P1、P2、P3区前体蛋白,P1区可进一步加工形成病毒衣壳蛋白VP1、VP2、VP3和VP4。P2和P3区主要编码非结构蛋白如2A、3C蛋白酶及3D RNA聚合酶,进化上具有高度保守性[11]。EV71的4种结构蛋白中,被广泛用于EV71基因分型的是VP1和VP4。VP4区核苷酸序列仅为207 bp,易扩增,但其高度保守,和VP1区相比核苷酸变异较少。而VP1位于病毒衣壳外侧,是目前EV71分型最常应用的区域。基于VP1序列的同源性差异,目前将EV71分为A、B、C基因型。其中,A基因型为EV71的原型株[12];B基因型可进一步分为B1~B5亚型,主要包括澳大利亚、新加坡部分地区、台湾省和马来西亚的病毒株;C基因型可划分为C1~C5亚型,主要是在亚太地区,包括中国大陆及台湾省、新加坡和马来西亚等地。
本研究采用RT-PCR方法分段扩增EV71南京市分离株NJ2017 iso1,2的基因组序列,并对其核苷酸序列进行了测定及分析,经序列拼接后得到病毒全基因组序列。本文主要对EV71的5′UTR区的核苷酸序列,结构蛋白VP1和VP4以及非结构蛋白2A、3C、3D的核苷酸和氨基酸序列进行同源性分析。经同源性比对发现,NJ2017 iso1,2之间全基因核苷酸序列和氨基酸序列同源性分别为94.0%和90.1%,VP1、VP4、2A、3C蛋白的核苷酸序列和氨基酸序列也具有高度同源性,但5′UTR区(25.6%)和3D蛋白(62.4%)的核苷酸序列差异较明显。本研究将NJ2017 iso1,2株与下载自GenBank中EV71流行株的核苷酸和氨基酸序列分别进行同源性分析,选择NJ2017 iso2作为参照。分析显示,NJ2017 iso2与A基因型和B基因型病毒株在VP1区的核苷酸序列差异均在15%以上,而与C基因型毒株在VP1区核苷酸序列差异均低于12%,其中与C4亚型各毒株的同源性较高,表明南京分离株均属于C4亚型毒株。经序列比对发现,NJ2017 iso2与2008年安徽阜阳分离株Fuyang 17.08-02(96.9%)以及2008年北京分离株BJ08-Z020-1(96.7%)的核苷酸同源性最高,与EV71原型株BrCr(79.8%)的同源性最低。从同源性分析表中可以直观地发现,虽然南京市分离株与其他地区EV71分离株之间核苷酸序列差异显著,但其氨基酸序列变化相对稳定。
2A、3C蛋白不仅作为病毒自身合成的蛋白裂解酶,在EV71的固有免疫逃避中也发挥着关键作用[13]。2A,3C 蛋白的核苷酸和氨基酸同源性分析可以看出,南京市 2 株 EV71 分离株之间同源性很高,这为病毒的体外感染和逃避宿主免疫起到了重要的作用。本实验中 2 株分离株的 3D 区核苷酸序列比对在所有区域中差异最大,这与Jiang等[14]和 Li 等[15]的研究结果相一致。3D蛋白可启动正链RNA的复制和VPg尿苷酸化的起始及终止[16],3D区核苷酸序列部分碱基的突变为病毒的进化提供了条件,同时自然选择压力又使病毒蛋白保持着较高的稳定性。同源性分析表中还可以发现,EV71南京市分离株之间的5′UTR区核苷酸序列差异性显著。5′UTR通过自身折叠形成的多个特异性结构与宿主的相关细胞蛋白结合,在起始病毒基因组RNA的合成以及病毒蛋白翻译的过程中发挥重要作用[17]。此外,5′UTR区还涉及病毒的毒力等方面的功能。南京市分离株之间5′UTR区核苷酸的显著性差异可能对其致病性产生的影响还需要进一步的研究。
基于VP1区构建的系统进化树显示,南京市分离株NJ2017与C4a基因亚型毒株聚成一簇,因此我们判断其为C4a亚型,这符合中国大陆地区近年来EV71病毒暴发的流行趋势[18],也说明南京市2017年EV71分离株没有出现明显的亚型变异。从进化树上可知,南京市分离株与上海、浙江等地的病毒分离株有较高的亲缘关系,同属于C4a基因亚型,可能是由于南京往返于上海、浙江等地的人员较为频繁,为病毒的快速传播提供了条件。然而EV71全基因组中结构蛋白基因仅占35%,而非结构蛋白则超过55%,肠道病毒之间的同源重组位点多位于非结构编码区[19],因此有研究认为仅以VP1区进行基因分型及进化分析会忽略 EV71 其他基因突变的信息,不能提供完整的种系发生关系[20],因此应参考 EV71 的全基因组序列对毒株完成基因分型及进化分析[21]。通过对全基因组的进化分析显示,A、B、C型分别位于3个大的分支中,与VP1区构建的进化树结构大体相似,但各基因亚型所在的分支并不完全相同,可能是不同的亚型之间存在着其他的变异信息。从两株进化树中可以看出,基于VP1基因序列分型的C4a亚型Guangdong 2009株在全基因组水平构建的进化树中被C4b亚型毒株分隔开以及B4亚型SB2864-SAR-00株单独成枝,说明仅依据VP1基因序列对EV71毒株进行基因分型是不能提供完整的种系进化信息。
近年来中国内地各地区EV71的暴发常以C4基因型为主,在病毒感染的早期阶段多为隐性感染,这为HFMD疫情的防控增加了难度。本研究通过对南京市EV71感染引发的HFMD患者的粪便样品进行了EV71病毒的分离并完成了病毒全基因组序列的测定及基因同源性和遗传进化分析,结果显示本研究中2017年南京市分离株均为C4a基因型,研究结果为南京市EV71的流行趋势研究和防控工作提供了参考依据。目前EV71的发病率仍呈上升趋势,但相应疫苗的研发已进入了临床阶段[22-23],相信不久的将来肠道病毒71型也会同脊髓灰质炎病毒一样被完全治愈。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
肝博士(2022年3期)2022-06-30 02:48:28
科学大观园(2022年2期)2022-01-23 11:05:15
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
中国病理生理杂志(2015年8期)2015-12-21 12:38:10
动物医学进展(2015年10期)2015-12-07 05:46:18
中国当代医药(2015年30期)2015-03-01 02:08:19
特产研究(2014年4期)2014-04-10 12:54:12
癌变·畸变·突变(2014年2期)2014-03-01 04:39:42
癌变·畸变·突变(2014年2期)2014-03-01 04:39:41
