
2种菊科植物对镉胁迫的生长和生理响应
2019-01-19 06:17王利芬孔丛玉吴思琳
江苏农业科学 2019年22期
王利芬 孔丛玉 吴思琳


摘要:以金盏菊(Calendula officinalis)、万寿菊(Tageteserecta)为材料,研究水培条件下0.0、0.5、1.0、2.0、4.0 mg/L不同质量浓度镉(Cd)胁迫对这2种菊科植物株高、总根长度、总根表面积、总根投影面积、根尖数及叶片中丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性等指标的影响。结果表明,0.5、1.0 mg/L 低质量浓度Cd胁迫处理下的万寿菊、金盏菊,其株高增长量高于0.0 mg/L(对照)处理,2.0、4.0 mg/L 高质量浓度Cd胁迫处理下的万寿菊、金盏菊,其株高增长量低于对照;随Cd胁迫质量浓度的增加,2种菊科植物叶片中MDA含量及金盏菊POD、CAT活性呈先增后减趋势,万寿菊叶片中SOD、POD、CAT活性及金盏菊CAT活性呈下降趋势,Cd胁迫质量浓度为0.5 mg/L时2种菊科植物叶片MDA含量高于对照;Cd胁迫质量浓度1.0 mg/L处理的万寿菊、2.0 mg/L处理的金盏菊其总根长度、总根表面积、总根投影面积、根尖数根系等生长指标相对较高。金盏菊和万寿菊对镉的耐性生理响应存在差异,金盏菊对Cd的耐性强于万寿菊。
关键词:花卉;菊科;镉;胁迫;耐性;生理特性
中图分类号:S682.1+10.1 文献标志码: A 文章编号:1002-1302(2019)22-0164-03
随着社会发展,各类污染问题随之而来,重金属污染便是其中之一。镉(Cd)作为一种污染范围及生物毒性很大的重金属,不仅会对土壤造成危害,且Cd随着工农业废水排出时,会给水体带来极大的污染,同时,由于Cd能够被植物吸收而进入食物链,因此其危害不仅仅是对水土和植物而言的,对动物、人类甚至各大生态系统都会带来严重影响[1],如何有效合理修复Cd污染问题亟待解决。在治理重金属污染的众多方法中,相较于物理、化学方法,利用植物进行修复不仅能够改善污染问题,还能美化环境,而且成本低廉、操作简单、不会破坏土壤结构,这使得植物修复受到越来越多环保者的青睐[2]。Cd会对植物产生许多毒害作用,如抑制根系活力,降低叶绿素含量和叶绿体数量,降低碳同化能力和新陈代谢速率等[3-6],筛选出耐Cd植物对利用植物修复Cd污染环境极为重要。
金盏菊(Calendula officinalis)、万寿菊(Tageteserecta)是园林绿化中常用的2种菊科草本观赏花卉,花期长、株型紧密、易栽培、好管理、适应性强,在生产中得到广泛应用。水培是一种利用营养液栽培植物的方式,可人为控制营养液中的养分、温度、溶解氧、酸碱度等环境条件[7],相比传统土壤栽培而言,水培营养吸收有效性高,养分供应充分、迅速,在相关科研和生产中被得到广泛应用[8-9]。本研究通过水培方式,探讨营养液中添加不同浓度Cd对万寿菊、金盏菊生长、抗氧化酶活性及根系生长等生理特性的影响,分析万寿菊和金盏菊在水培条件下对Cd胁迫的生理响应机制,为其在Cd污染修复中的应用提供理论依据。
1 材料与方法
1.1 试验处理
供试万寿菊品种为“发现”、金盏菊品种为“棒棒”,其种子均来源于虹越花卉股份有限公司。2017年11月上旬进行播种,挑选大小规格均一、饱满的种子,经催芽后播种在海绵块中;将海绵块放入水培盆中,置于培养室内进行培养,温度保持在26~28 ℃,光照度为5 000~6 000 lx,光照时间为 12 h/d,培养液采用霍格兰营养液;当幼苗长出3~4张真叶时,挑选生长状况基本一致的幼苗移栽到含Cd(试剂为 CdCl2·2.5H2O)质量浓度分别为0.5、1.0、2.0、4.0 mg/L的不同营养液上进行水培,以Cd质量浓度为0.0 mg/L处理为对照(CK),每处理5株幼苗,重复3次。
1.2 测定指标及方法
1.2.1 株高 移栽时及之后每隔7 d,采用数显游标卡尺测量株高,直至移栽28 d结束,植株株高增长量为培养28 d时植株株高与移栽时株高的差值。
1.2.2 根系生长指标 镉胁迫处理28 d,每处理随机选取植株3株,采用爱普生中国股份有限公司产Epson Perfection V700 Photo型扫描仪对其根系进行扫描,利用根系分析系统WinRHIZO得出总根长度、总根表面积、总根投影面积、根尖数。
1.2.3 丙二醛含量和保护酶活性 镉胁迫处理28 d,选取同一部位植株叶片,分别采用硫代巴比妥酸显色法、氮蓝四唑还原法、愈创木酚显色法、紫外分光光度法测定丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性[10-12]。
1.3 数据分析
采用Excel 2010软件对数据进行整理统计及图表制作。
2 结果与分析
2.1 Cd胁迫对2种菊科植物株高增長量的影响
由图1可见,Cd胁迫处理下,金盏菊、万寿菊植株的生长受到不同程度影响,随Cd胁迫浓度的增加,2种菊科植物株高增长量呈先增后减趋势,0.5、1.0 mg/L低质量浓度Cd处理的2种菊科植物株高增长量都略高于对照,2.0、4.0 mg/L高质量浓度Cd处理的2种菊科植物株高增长量都略低于对照,低质量浓度镉胁迫可促进植株的生长,而高质量浓度抑制植株的生长;同一Cd质量浓度胁迫处理下,金盏菊株高增长量明显高于万寿菊。
2.2 Cd胁迫对2种菊科植物根系生长的影响
由表1可见,随Cd胁迫质量浓度的增加,2种菊科植物总根长、总根投影面积、总根表面积、根尖数等根系生长指标总体呈先增后减趋势;镉胁迫质量浓度为1.0 mg/L时,万寿菊的总根长、总根投影面积、总根表面积、根尖数相对最大,分别比对照增加68.56%、24.35%、35.52%、75.51%,而镉胁迫质量浓度为2.0 mg/L时,金盏菊的总根长、总根投影面积、总根表面积、根尖数相对最大,分别比对照增加37.44%、11.80%、25.06%、35.92%。
2.3 Cd胁迫对2种菊科植物叶片MDA含量保护酶活性的影响
2.3.1 MDA含量 由图2可见,随Cd胁迫质量浓度的增加,2种菊科植物叶片中MDA含量呈先增后减趋势;Cd胁迫质量浓度为0.5、1.0 mg/L时,万寿菊叶片MDA含量高于对照,且在Cd质量浓度为0.5 mg/L时MDA含量相对最高,为对照植株的 2.83倍,Cd胁迫质量浓度为2.0、4.0 mg/L时,万寿菊叶片中MDA含量低于对照;Cd胁迫对金盏菊叶片MDA含量的影响没有万寿菊明显,Cd胁迫质量浓度为 0.5 mg/L 时,金盏菊叶片中MDA含量高于对照,其他质量浓度胁迫处理均略低于对照。
2.3.2 SOD活性 由图3可见,随Cd胁迫质量浓度的增加,万寿菊叶片中SOD活性呈下降趋势,Cd胁迫质量浓度为4.0 mg/L时,其SOD活性相对最低,仅为5.40 U/g;与万寿菊不同,随Cd胁迫质量浓度的增加,金盏菊叶片中SOD活性呈先增后降趋势,Cd胁迫质量浓度为0.5 mg/L时,金盏菊叶片中SOD活性相对最高,略高于对照,而其他Cd质量浓度胁迫处理均低于对照,且下降幅度较小。
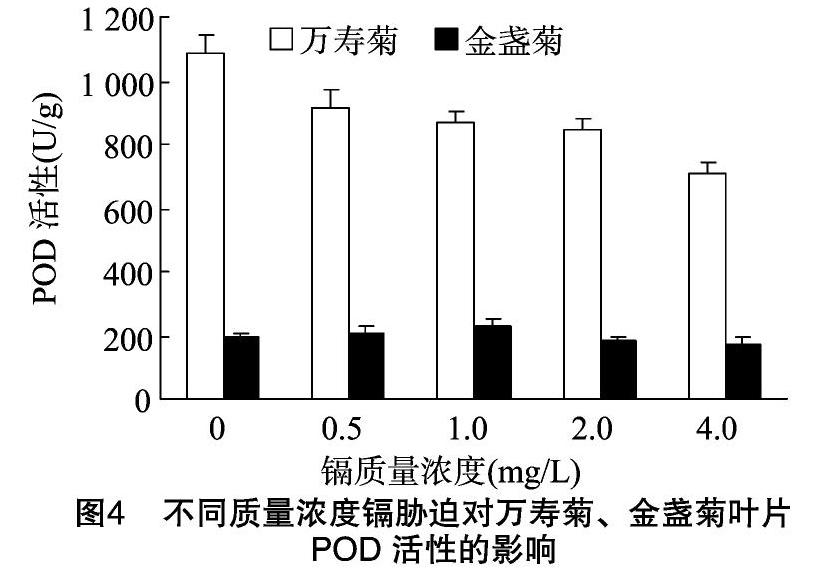
2.3.3 POD活性 过氧化物酶是植物产生的一类氧化还原酶,与呼吸作用、光合作用等许多植物生长发育过程中的重要反应相关,其酶活性高低与植物代谢强度有密切关系,可使生物活性氧维持在一个較低水平上,以防止活性氧的伤害[13]。由图4可见,随Cd胁迫质量浓度的增加,万寿菊叶片中POD活性呈下降趋势,POD活性由对照的1 087.50 U/g下降到Cd胁迫质量浓度为4.0 mg/L处理时的710.14 U/g,下降幅度为 34.70%,这与其SOD活性规律相似;随Cd胁迫质量浓度的增加,金盏菊叶片中POD活性呈先增后降趋势,与对照相比,总体增减幅度相对较小,POD活性维持在224.07~168.67 U/g,Cd胁迫质量浓度为0.5、1.0 mg/L时金盏菊叶片POD活性略高于对照,2.0、4.0 mg/L Cd质量浓度处理的金盏菊叶片POD活性略低于对照。
2.3.4 CAT活性 由图5可见,随Cd胁迫质量浓度的增加,万寿菊叶片中CAT活性呈明显下降趋势,对照叶片中CAT活性相对最高,为82.15 U/g,Cd胁迫质量浓度为 4.0 mg/L 时CAT活性相对最低,仅为13.48 U/g,为对照的16.41%;金盏菊叶片中CAT活性变化规律与万寿菊相似,但其CAT活性值明显低于万寿菊;Cd胁迫质量浓度为0.5、1.0 mg/L 时,金盏菊叶片中CAT活性略低于对照,Cd胁迫质量浓度增加到2.0 mg/L时,金盏菊叶片中CAT活性较对照下降明显,Cd胁迫质量浓度达到 4.0 mg/L 时,其CAT活性仅为0.46 U/g。
3 结论与讨论
镉作为一种非必需元素,可能与植物必需营养元素竞争根部的吸收位点,或通过影响植物生理生化过程来影响植物的正常生长,从而在高质量浓度下造成植物生物量的下降[14]。本试验结果表明,随Cd胁迫质量浓度的增加,万寿菊、金盏菊植株高度增长量呈先增后降的规律,0.5 mg/L Cd质量浓度处理的万寿菊、1.0 mg/L Cd质量浓度处理的金盏菊,其植株株高增长量相对最大;从根系生长状况看,低质量浓度Cd胁迫可以促进万寿菊、金盏菊这2种菊科植物根系的生长,根系总根长、总投影面积、总表面积、根尖数的最大值出现时万寿菊Cd胁迫质量浓度为1.0 mg/L,而金盏菊则为 2.0 mg/L。这在一定程度上反映根系对Cd的耐受质量浓度高于植株地上部分,金盏菊对Cd的耐受能力强于万寿菊。
重金属胁迫等不良环境能诱发植物代谢过程产生自由基,从而对植物膜产生伤害作用,而植物自身的保护酶系统能够清除自由基,以减轻其危害[15]。超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)是植物体内活性氧清除系统的组成部分,能有效清除植物体内的自由基和过氧化物[16]。本试验结果表明,随Cd胁迫质量浓度的增加,金盏菊叶片中POD、CAT活性呈先增后减趋势,万寿菊叶片中SOD、POD、CAT活性呈下降趋势,且高质量浓度Cd胁迫下,POD、CAT活性下降明显,这可能是由于植物体内产生的活性氧不能被及时清除而质子化成毒性更强的·OH自由基有关,这与田治国等的研究结论[17]较为吻合。
万寿菊、金盏菊这2种菊科植物对Cd毒害响应存在明显的品种差异,对Cd的耐受质量浓度有所不同,相对而言,金盏菊对Cd胁迫具有较高的耐性,可作为绿化植物在城市镉污染水体或土壤严重的地区种植,以用于对土壤或水体的修复。
参考文献:
[1]Wang L,Zhou Q X,Ding L L,et al. Effect of cadmium toxicity on nitrogen metabolism in leaves of Solanum nigrum L. as a newly found cadmium hyperaccumulator[J]. Journal of Hazardous Materials,2008,154(1/3):818-825.
[2]苗欣宇,周启星. 污染土壤植物修复效率影响因素研究进展[J]. 生态学杂志,2015,34(3):870-877.
[3]何俊瑜,王阳阳,任艳芳,等. 镉胁迫对不同水稻品种幼苗根系形态和生理特性的影响[J]. 生态环境学报,2009,18(5):1863-1868.
[4]Baryla A,Carrier P,Franck F,et al. Leaf chlorosis in oilseed rape plants (Brassica napus) grown on cadmium-polluted soil:causes and consequences for photosynthesis and growth[J]. Planta,2001,212(5/6):696-709.
[5]张金彪,黄维南. 镉胁迫对草莓光合的影响[J]. 应用生态学报,2007,18(7):1673-1676.
[6]万雪琴,张 帆,夏新莉,等. 镉处理对杨树光合作用及叶绿素荧光参数的影响[J]. 林业科学,2008,44(6):73-78.
[7]洪坚平,谢英荷,孟会生,等. 水培油菜营养液养分动态变化研究[J]. 中国农学通报,2008,24(1):330-334.
[8]曹晨书,曾春霞. 蔬菜水培技术的研究进展[J]. 上海蔬菜,2012(6):3-4.
[9]张启翔,康红梅,唐 菁,等. 切花月季无土栽培技术的研究[J]. 北京林业大学学报,2003,25(3):22-27.
[10]郝再彬,苍 晶,徐 仲. 植物生理实验[M]. 哈尔滨:哈尔滨工业大学出版社,2004:115-116.
[11]曲 敏,秦丽楠,刘羽佳,等. 两种检测SOD酶活性方法的比较[J]. 食品安全质量检测学报,2014(10):3318-3323.
[12]李仕飞,刘世同,周建平,等. 分光光度法测定植物过氧化氢酶活性的研究[J]. 安徽农学通报,2007,13(2):72-73.
[13]吴建勋,张姗姗. Cr、Co、Pb单一胁迫对浮萍SOD、POD、MDA的影响[J]. 中国农学通报,2013,29(15):188-194.
[14]涂 从,郑春荣,陈怀满. 土壤-植物系统中重金属与养分元素交互作用[J]. 中国环境科学,1997,17(6):526-529.
[15]蒋兴一,李景吉,钱美玲. 三种菊科草本植物对重金属Cd污染土壤的修复效应实验研究[J]. 四川环境,2017,36(5):29-33.
[16]奉 斌,代其林,王 劲. 非生物胁迫下植物体内活性氧清除酶系统的研究进展[J]. 绵阳师范学院学报,2009,28(11):50-53,77.
[17]田治国,王 飞. 不同品种万寿菊对镉胁迫的生长和生理响应[J]. 西北植物学报,2013,33(10):2057-2064.
猜你喜欢
天然产物研究与开发(2018年10期)2018-11-06
山东农业科学(2016年12期)2017-01-21
现代园艺(2016年9期)2016-07-16
当代化工研究(2016年5期)2016-03-20
现代园艺(2016年2期)2016-03-15
绿色科技(2015年6期)2015-08-05
山东农业科学(2014年8期)2014-10-10
