
种源地气候对蒙古莸光响应特性的影响
2019-01-18 05:51白雪卡纪若璇王襄平
生态学报 2018年23期
白雪卡,刘 超,纪若璇,沈 超,王襄平,*
1 北京林业大学, 林学院森林资源与生态系统过程北京市重点实验室, 北京 100083 2 北京林业大学, 生物科学与技术学院林木育种国家工程实验室, 北京 100083
植物的光合作用是决定群落生产力大小的重要的生理生态过程[1]。植物对光强的响应一直是植物生理生态学研究中的热点问题[2- 4]。光响应曲线是研究光强与净光合速率关系的模型,通过分析光响应曲线可以得到植物的最大净光合速率、光饱和点、光补偿点、暗呼吸速率等重要的光合参数[5- 6],由此可以进一步了解植物的光化学过程及其对光合有效辐射的利用能力[7]。
同一物种长期在不同的环境中为了适应其特定的地理环境,会在表型和基因等方面产生一定程度的变异[8- 10],进而形成不同的种源特性。这是植物在自然选择条件下的一种适应,这种适应对同一物种造成的改变,称之为种内遗传变异[11],造成这种遗传变异的主要因素是遗传和环境[12- 13]。由种源地环境的差异造成的种源变异主要体现:(1)表型可塑性(不同环境下基因表达不同);(2)基因型差异(不同环境导致的基因型差异);(3)两者共同造成的。在不同环境下,植物叶片的光合等性状会因种源地温度、水分等环境气候因子的差异而不同[14],对植物光合能力存在着不同程度的影响[15- 16];而把不同种源的植物移植到同一生长环境下,相同的生长环境条件排除了环境梯度的影响,不同种源植物叶性状表现出差异是由于原种源地环境差异造成的植物遗传性基因变异,属于遗传因素。关于光合特征等叶性状随气候梯度的变化,我国学者已经开展一些重要的研究[17- 18]。不过,这些研究是测定的是不同气候条件下的植物,其光合等特征受到环境因素和遗传因素的共同影响(包括气候梯度和种源的影响)。开展在相同环境条件下不同种源的光合特征的研究,有助于在上述研究基础上,进一步分析种源地的气候条件如何影响植物的光合,从而深入理解光合作用的影响因素。
蒙古莸(Caryopterismongholica)是唇形科(Lamiaceae)莸属(CaryopterisBunge)的抗旱性灌木[19- 20],常广泛分布于内蒙古、甘肃、宁夏等沙丘荒野、干涸河床、砾石山地,是一种耐旱、耐寒兼具观赏性的固沙绿化灌木[21- 23]。韩磊等[24]研究蒙古莸的叶水势与环境因素的关系,发现蒙古莸叶水势日变化与气温、光辐射强度、大气相对湿度及土壤含水量显著相关。谢乾瑾等[25]发现,干旱胁迫下蒙古莸光合能力的下降是引起其生长减缓的重要原因。目前,研究学者对植物光合作用的研究已经不再局限于光合速率与光强、小尺度环境因子的关系,而是研究大尺度的环境因素和遗传因素如何共同影响植物光合作用,二者的相对作用大小如何等问题[26- 28]。
为了更好的研究遗传因素对植物光合作用的影响机制,我们将8个不同种源蒙古莸种植在同一环境条件下,排除外界环境因素差异的作用,对其光合作用光响应曲线特性进行研究,探讨以下问题:(1)不同种源蒙古莸之间的光合作用特性的差异性;(2)种源地气候对蒙古莸光合作用的影响作用大小。以期为今后蒙古莸的研究,以及由环境差异造成的植物基因型差异对植物光合作用机制的影响提供理论依据。
1 材料和方法
1.1 试验地概况
试验地点为北京林业大学苗圃(40.01N°,116.34E°),气候属于暖温带半湿润大陆性季风气候,年均降水量在500—650 mm之间,降水集中在7—9月,占全年降水量70%;年均温为11.8℃,全年最高温度为37.5—42.6℃,最低温度为-19.5—-14.8℃,7月温度约为25.8℃。
1.2 实验材料
试验材料为蒙古莸1年生扦插苗,种源来源及地理信息见表1。
于2016年3月下旬,在内蒙古、甘肃、宁夏等地采取生长良好蒙古莸植株,剪取长度约为20 cm左右的枝条茎段,放于湿布中保湿,带回北京。于4月上旬,将不同种源地的扦插枝条经0.1% 生根粉ABT1(中国林业科学研究院林业研究所)处理后扦插于450 mm×450 mm的花盆中,每个种源扦插苗木6盆,每盆3株,共18株。种植土壤为沙壤土(沙子∶壤土∶草炭土=45∶60∶60),与蒙古莸的野外土壤条件相似,田间持水量19.11%,土壤容重1.273 g/cm3。
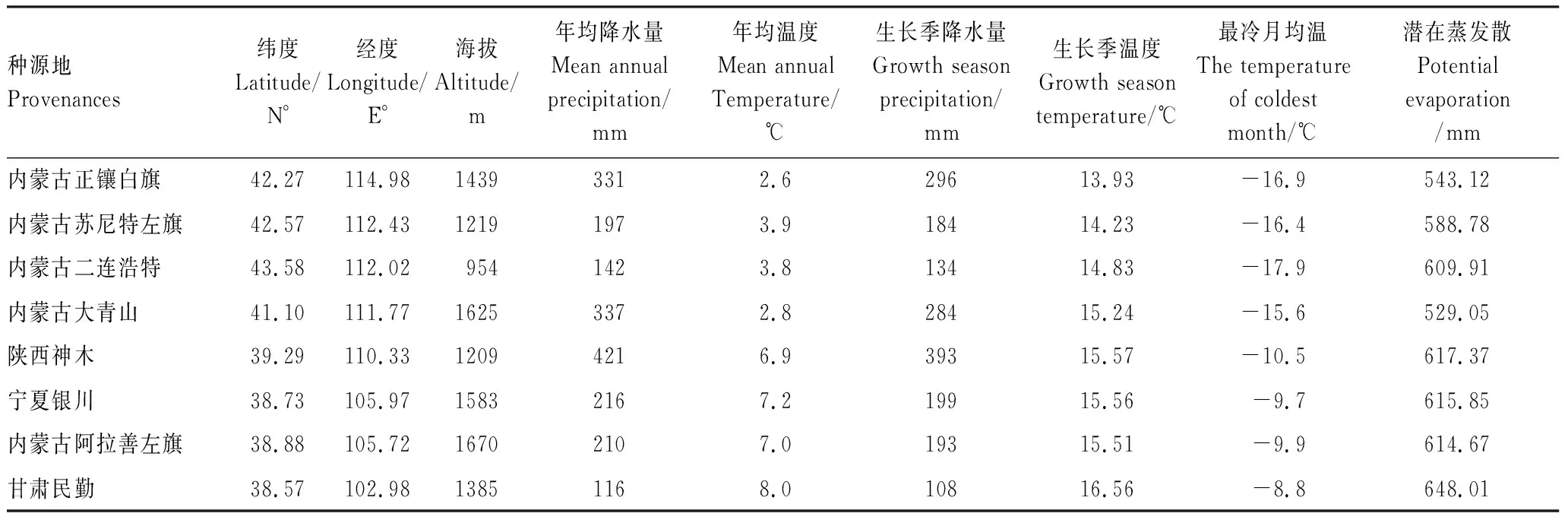
表1 蒙古莸种源地基本情况
待苗木正常生长3个月后,于7月中旬选取生长良好、一致的苗木进行试验,试验时将每盆水分控制在田间持水量的70%—80%。
1.3 测定指标和方法
为了使本处理稳定在试验设定土壤含水量内,采用烘干和称重的方法测定土壤含水量,试验时每天18:00按照预先模拟盆重与土壤含水量的关系,根据其变化补充消耗的水分。
实验时期选择2016年7月中旬连续晴天的上午(9:00—11:30)进行,此时段大气平均气温约为(32±2)℃,光照强度约为(800±100)μmol m-2s-1,处于短期高温、高光的环境条件。
每个种源随机选择3株健康植株,并选取成熟的向阳叶片进行测定,叶位为自上而下第7—8片叶。采用Li-Cor 6400便携式光合作用测定系统(Li-Cor,USA)进行光响应曲线的测定,光源采用Li- 6400- 40B光源,CO2含量为(360±2) μmol/mol,流速为500 mmol/s,叶片温度为(26±1)℃,空气相对湿度约为70%。光合有效辐射梯度设置为: 2000、1800、1600、1400、1200、1000、800、600、400、200、100、50、20、10、0 μmol m-2s-1。采用Ye等[29- 30]的直角双曲线修正模型进行曲线拟合,得到表观量子效率(α1),最大净光合速率(Pnmax),光饱和点(Isat)、光补偿点(Ic)和呼吸效率(RE)等光合参数。
直角双曲线修正模型表达式:
(1)
式中,α是光响应曲线的初始斜率,也称为初始量子效率,β和γ为系数,I为光合有效辐射,Rd为暗呼吸速率。
最大净光合速率(Pnmax)为:

(2)
光饱和点(Isat)为:
(3)
光补偿点(Ic)为:
(4)
表观量子效率(α1)为:
α1=Rd/Ic
(5)
式中,α1是光响应曲线在光补偿点(Ic)处的斜率。
呼吸效率(RE)为:
RE=Pnmax/Rd
(6)
根据蒙古莸各个种源地的经纬度,从ArcGIS 10.2中WorldCLIM 全球高分辨率气候据库[31]中提取出各样地的50年月平均气候数据(1950—2000),计算各个气候因子大小。包括年均温(MAT);年均降水量(MAP):反映水分供给情况;生长季降水量(GSP):4月到10月的降水量;生长季温度(GST):4月到10月的平均温度;最冷月均温(MTCM):1月份的温度,反映冬季低温;潜在蒸发散(PET):反映年总热量供给。
1.4 数据分析
用方差分析多重比较(ANOVA-Duncan)方法,比较不同种源蒙古莸光响应参数的差异。用Pearson相关性分析种源地环境因子与各个光合参数的相关关系;并结合一般线性模型(general linear model,GLM)ANOVA分析种源地气候对蒙古莸各个光合参数值的影响作用大小[32]。
一般线性模型ANOVA分析中气候因子选取原则:
1)基于种源地环境因素之间的相关性(表3),及其与各个光合参数值的相关关系(表4),选取代表种源地水分、温度和光三大元素的4个气候因子:生长季降水(GSP)、生长季温度(GST)、最冷月均温(MTCM)和海拔(Altitude);
2)采用BIC(Bayesian Information Criterion)模型对上述4个种源地气候因子进行筛选,得到最终进入一般线性模型ANOVA分析中的气候因子。
数据分析在Excel和R软件(R3.2.2)中完成。
2 结果
2.1 不同种源蒙古莸光合参数的差异
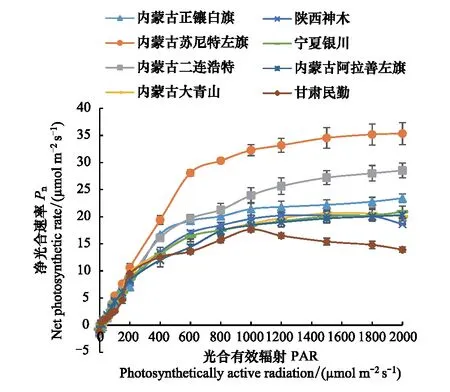
图1 不同种源蒙古莸的净光合速率对光强的响应曲线 Fig.1 The response curve of the net photosynthetic rate to the light of different Caryopteris mongholica provenances*图中误差线代表平均值的标准误差
由图1和表2可以得到,各个种源的蒙古莸光合作用光响应曲线具有明显差异,且波动范围较大。从图1中看,内蒙古苏尼特左旗种源和内蒙古二连浩特种源的蒙古莸最大净光合速率(Pnmax)较高,甘肃民勤种源的较低。
表观量子效率(α1)是光合过程中指示光能转化效率的指标之一[33],由表2可以看出,不同种源蒙古莸光能转化效率有着差异,α1的范围为:0.0533—0.0754。甘肃民勤种源的表观量子效率(α1)最小,内蒙古苏尼特左旗种源的α1最大,且差异显著,说明内蒙古苏尼特左旗的蒙古莸光能转化率高。
最大净光合速率(Pnmax)是植物最大光合速率与呼吸速率的净值,它代表植物光合作用的能力,反映了植物最大光合潜力[30],这8个种源的Pnmax范围为:16.69—35.67 μmol m-2s-1,其中内蒙古苏尼特左旗种源的Pnmax最大,甘肃民勤种源的Pnmax最小。内蒙古苏尼特左旗、二连浩特与甘肃民勤种源的Pnmax差异显著(P<0.05),其余5个种源Pnmax差异不显著。
光饱和点(Isat)是衡量植物利用强光能力的一个指标,蒙古莸的Isat的范围为:1122.05—2939.01 μmol m-2s-1,波动范围较大,其中内蒙古二连浩特种源Isat最大,甘肃民勤种源的最小,大多数种源的Isat在1800 μmol m-2s-1左右,内蒙古苏尼特左旗和大青山种源的Isat差异不显著,说明这2个种源利用强光的能力较为接近。
光补偿点(Ic)是指示植物利用弱光能力的一个指标,Ic越高,说明植物利用弱光的能力越弱[33]。内蒙古阿拉善左旗种源Ic(8.19 μmol m-2s-1)最低,该种源利用弱光的能力强;内蒙古正镶白旗种源Ic(28.83 μmol m-2s-1)最大,其中内蒙古苏尼特左旗、陕西神木与甘肃民勤种源Ic差异不显著,说明这3个种源对弱光的利用能力相似。
暗呼吸速率(Rd)表示植物正常生理活动中提供的必须能量,大多种源Rd范围在1—1.5 mol m-2s-1左右。Rd最小的内蒙古阿拉善左旗种源(0.46 mol m-2s-1),最大的是内蒙古正镶白旗种源(1.92 mol m-2s-1),说明内蒙古正镶白旗种源消耗的能量较多,有机质积累较少。
呼吸效率(RE)表示最大净光合速率与暗呼吸速率的比值,呼吸是植物重要的生理过程之一。这8个种源的RE差异较大,其中RE最大的是内蒙古阿拉善左旗种源46.22 mol m-2s-1,最小的是内蒙古正镶白旗种源12.15 mol m-2s-1,内蒙古二连浩特、大青山种源、陕西神木种源和甘肃民勤种源RE大致在20 mol m-2s-1左右,且差异不显著,说明这4个种源的呼吸效率差异不大。
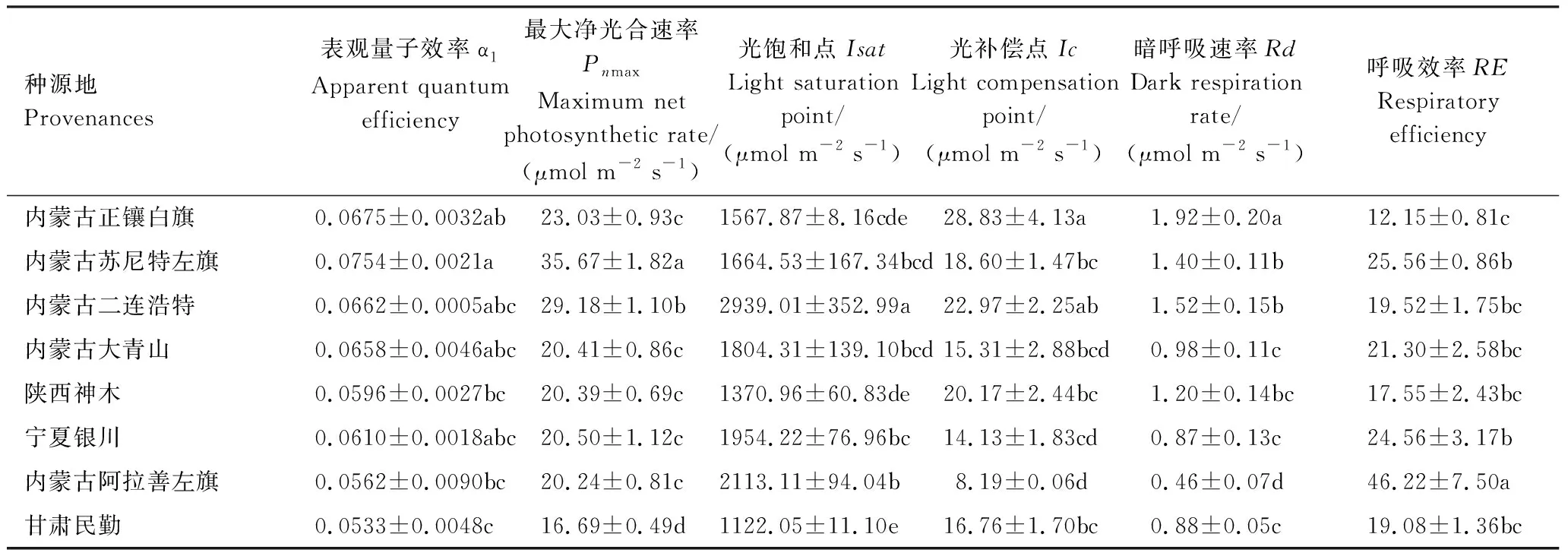
表2 由光响应曲线所得的光合参数值
* 表中数值为平均值±标准误差; 表中字母为各个光合参数的一维方差分析进行的Duncan多重比较检验分组,同一光合参数的不同字母表示差异性显著(P<0.05)
2.2 种源地气候对蒙古莸光合参数的影响
由表3可得,气候指标间存在明显的自相关,且温度与经纬度密切相关。其中生长季温度(GST)与年均温度(MAT)、最冷月均温(MTCM)相关性极显著(P<0.001);生长季降水(GSP)与年均降水量(MAP)相关性也极显著;海拔(Alititude)与纬度(Latitude)相关性显著(P<0.01),与其他气候因子相关性均不显著。
由表4可以看出,Pnmax、Ic、Rd与经纬度显著正相关(P<0.05),与海拔显著负相关(P<0.05),说明随着经纬度的增加和海拔的降低,蒙古莸的光合能力逐渐增强。降水指标(MAP和GSP)与各个光合参数值的相关性均不显著;温度指标(MAT、GST和MTCM)与α1、Pnmax、Ic、Rd均显著(P<0.05)负相关;潜在蒸发散(PET)只与α1显著(P<0.05)负相关。在相同的环境条件下生长时,蒙古莸光合作用受种源地的地理位置(经纬度)、温度(MAT、GST和MTCM)和海拔的显著影响。

表3 环境之间的相关系数
* MAP: Mean annual precipitation; MAT: Mean annual temperature; PET: Potential evaporation; GSP: Growth season precipitation; GST: Growth season temperature; MTCM: The temperature of coldest month; ***P<0.001; **P<0.01; *P<0.05

表4 种源地环境因素与各个光合参数值的相关系数
*MAP: Mean annual precipitation; MAT: Mean annual temperature; GSP: Growth season precipitation; GST: Growth season temperature; MTCM: The temperature of coldest month; PET: Potential evaporation; ***P<0.001; **P<0.01; *P<0.05
通过一般线性模型(GLM)ANOVA分析(表5)可知,种源地气候作为遗传因素对蒙古莸各个光合参数均有显著作用,其解释程度在16.65%—73.52%之间。其中,对α1、Pnmax、Ic和Rd解释程度均大于40%,影响极显著(P<0.001),但对RE的解释程度最小(16.65%)。种源地气候因子中,生长季降水(GSP)对Pnmax和Isat有影响,但不显著;生长季温度(GST)对α1、Pnmax、Ic和Rd的解释程度为:26.66%—58.33%,影响均显著(P<0.01);最冷月均温(MTCM)解释程度为18.35%,且影响显著(P<0.05);海拔(Altitude)对Pnmax、Ic、Rd和RE的解释程度在11.75%—16.65%之间,影响显著(P<0.05)。温度对蒙古莸光响应参数的解释程度大于海拔的。
3 结论与讨论
光合作用对植物的生长发育具有重要的意义,而光是植物光合作用必不可少的因素[35],不同植物对光的响应不同,同一植物的不同种源对光的响应也不同[34, 36- 37]。本研究对8个不同种源的蒙古莸光响应曲线的研究发现,蒙古莸光合响应能力在不同种源间表现出明显的不同,光响应曲线参数也存在着显著的差异(表2),地理梯度上显示从西到东,从南到北,蒙古莸的光合能力逐渐增强,最东北部的内蒙古二连浩特和苏尼特左旗种源蒙古莸的Pnmax较大,最西部的甘肃民勤种源较小(表2、表4)。相关研究表明,同一植物不同种源为了生存,经过长期对各自生长环境的适应,形成了各自独特的光合作用机制[38- 39],即使当不同种源植物移植到相同环境下生存,其光合能力仍存在差异[10, 13, 40- 42]。这种不同种源间光响应特性的差异正是植物趋异适应的体现。钟培芳[13]、刘海燕[43]、安海龙[42]等对不同种源白刺、沙柳、黄柳光合速率等光合特性及水分利用效率的研究也发现这种光合趋异性。
表5种源地气候(生长季降水、生长季温度、最冷月均温、海拔)对各个光合参数的GLM分析
Table5Summaryofgenerallinearmodelsfortheeffectofprovenanceclimatevariations(GSP, GST, MTCM, Altitude)onindividualphotosyntheticcharacteristics
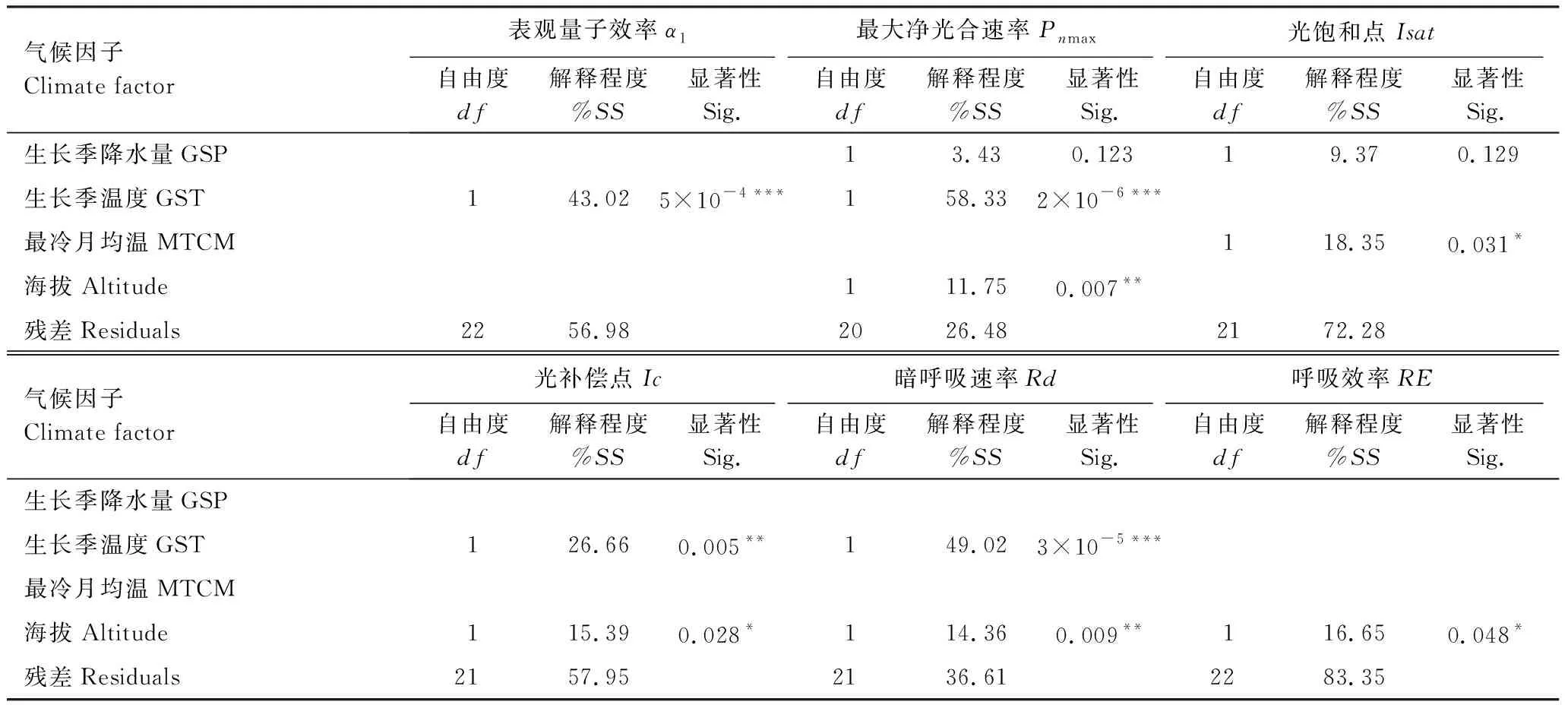
气候因子Climate factor表观量子效率α1最大净光合速率Pnmax光饱和点Isat自由度df解释程度%SS显著性Sig.自由度df解释程度%SS显著性Sig.自由度df解释程度%SS显著性Sig.生长季降水量GSP13.430.12319.370.129生长季温度GST143.025×10-4***158.332×10-6***最冷月均温MTCM118.350.031*海拔Altitude111.750.007**残差Residuals2256.982026.482172.28气候因子Climate factor光补偿点Ic暗呼吸速率Rd呼吸效率RE自由度df解释程度%SS显著性Sig.自由度df解释程度%SS显著性Sig.自由度df解释程度%SS显著性Sig.生长季降水量GSP生长季温度GST126.660.005**149.023×10-5***最冷月均温MTCM海拔Altitude115.390.028*114.360.009**116.650.048*残差Residuals2157.952136.612283.35
df,: degree of freedom; %SS: percentage of explained sum of squares; Sig.:significant; GSP: Growth season precipitation; GST: Growth season temperature; MTCM: The temperature of colddest month; ***P<0.001, **P<0.01, *P<0.05
造成不同种源植物之间光合能力趋异性的因素有很多,比如基因型变异、气候差异、群落结构不同、地理位置差异等[14- 16]。本研究将不同种源的蒙古莸种植在相同环境中,排除了生长环境的气候差异、群落差异等因素,主要研究由种源地气候差异所引起遗传变异对蒙古莸光合能力的影响。结果表明,种源地温度(GST、MTCM)与海拔(Altitude)对蒙古莸光合能力有着显著影响(表4、表5)。Sack等[44]报道称植物光合进化适应主要与温度决定的生长季长短、海拔决定的植物光胁迫等有关。安海龙等[42]在同一环境对不同种源黄柳(Salixgordejevii)叶性状的研究中,净光合速率等光合指标受种源地气候(MAT、MAP)的显著(P<0.05)影响。不同种源植物种植在同质园或相同环境下,种源地气候作为遗传因素显著影响着植物的叶性状[10, 12- 14]。
在同一环境下,蒙古莸的光合参数(α1、Pnmax、Ic和Rd)受种源地温度(MAT、GST、MTCM)的影响,随着温度的升高,其光合能力逐渐降低(表4);而Isat受低温影响较大,与最冷月均温(MTCM)显著负相关,与生长季温度(GST)没有显著相关性(表4、表5)。冯秋红[45]、Wright[46]等大尺度研究也有类似的结果。但钟培芳等[13]对4个种源白刺(Nitrariatangutorum)种植在同一环境中光合作用的研究,却发现其光合速率不受种源地温度的影响作用,而受种源地降水的影响较大。季子敬等[10]对不同种源兴安落叶松(Larixgmelinii)的研究中也可以得到,其最大净光合速率与种源地温度的相关性不显著。这可能是由于不同物种对温度的适应能力有所不同,进而导致其对种源地温度的响应有所不同[10, 13, 40]。也有研究指出,随着温度升高,植物光合能力下降,可能是植物比叶重、叶氮含量及光合氮素利用效率等的综合变化引起的[47- 48]。同时本研究中,种源地海拔与蒙古莸的Pnmax、Ic和Rd显著负相关,与RE显著正相关(表4)。而Soolanayakanahally等[49]研究中,在同质环境下,来自高海拔的种源树木为了长期适应较短生长季节,具有较高的净光合速率。由于蒙古莸的种源地分布在干旱半干旱地区,光照强度较强,高海拔地区种源为避免紫外线对细胞器的伤害,光合速率较低,会表现为较低的最大光合速率;在弱光下表现为海拔越高,光补偿点越低[44, 47, 50- 51]。
将种源地气候作为导致植物基因型变异的因素之一,本研究通过一般线性模型ANOVA分析发现,种源地气候对这6个光合参数解释程度在16.65%—73.52%之间,其中对α1、Pnmax、Ic和Rd这4个光响应参数的解释程度更是大于40%(表5)。研究将不同种源植物移植到相同环境下生存,排除了外界环境因素差异的作用,更为纯粹的反映了环境差异所引发的遗传变异效果。而在安海龙等[42]的类似分析中,种源地气候对黄柳光合作用有显著的解释力,而其解释程度相对降低(3.19%—13.77%)则是因为短期干旱处理较大影响了植物光合作用,导致水分胁迫(主因素)对其光合作用的解释程度较大(26.5%—76.81%)。
除此之外,造成不同种源蒙古莸光合能力差异的因素可能还有很多,CO2浓度、土壤养分含量等其他种源地环境因素造成的遗传差异也对植物光合能力具有一定影响[48, 52- 53],它们也可能造成种源间的遗传差异。种源地气候作为反映遗传因素的影响是造成不同种源蒙古莸光响应特性差异的重要因素之一,这为今后蒙古莸的引种栽培、种源选择及遗传资源保护提供了重要的理论依据。
猜你喜欢
特产研究(2022年6期)2023-01-17
科学(2022年4期)2022-10-25
干旱气象(2022年1期)2022-03-15
辽宁林业科技(2021年1期)2021-03-16
北方文学(2019年21期)2019-08-18
武夷学院学报(2017年9期)2017-11-29
小雪花·初中高分作文(2017年4期)2017-06-29
Coco薇(2016年5期)2016-06-03
西南农业学报(2016年5期)2016-05-17
广西林业科学(2016年3期)2016-03-16
