
高温对葡萄休眠芽萌发及需热量的影响
2019-01-18 02:50罗国安张亚红耿晓玲
西南农业学报 2018年12期
罗国安,张亚红*,李 鹏,耿晓玲
(1.宁夏大学农学院,宁夏 银川 750021;2.银川大学,宁夏 银川 750021)
【研究意义】植物休眠(Endormancy)是落叶果树生长发育过程中的生理现象,体现其对环境及季节性变化的适应性[1]。解除休眠是落叶果树由休眠到生长的恢复,需要一定的低温积累[2],而低温积累不足会导致落叶果树不能完整的结束休眠,会导致芽萌发延迟、萌芽率低、芽的萌发不整齐等[3-6],高温胁迫打破休眠的研究能为落叶果树促早栽培提供更多的选择。【前人研究进展】关于高温打破落叶果树芽休眠的研究如下:Wample等[7]在1993年试验发现52~60 ℃处理Cabernet Sauvignon葡萄休眠芽10~30 min延迟了芽的萌发,且58~60 ℃处理30 min让芽的存活率将为0 %;王海波[8]发现在高温条能够打破桃芽休眠,进而使其提前萌发。葡萄的需热量(heat requirement)是指从自然休眠结束至其萌芽展叶50 %所需的有效热量累积,又称热量单位积累或蓄热积温[9]。葡萄在结束生理休眠之后,需要一定的热量积累才能萌芽展叶。在葡萄实际生产工作中,需热量及其重要,它影响着果树萌芽展叶的速率。在过去几十年里,国内外对落叶果树需热量的研究主要集中在不同品种果树需热量数值、需热量与需冷量的关系研究上,如Okie WR[10]等的研究表明高需冷量品种桃同时具有高需热量,部分低需冷量品种同时具有低需热量,需热量和需冷量表现出互补关系;王海波等[11]的研究发现葡萄的需冷量和需热量之间呈负相关关系。关于外界因素如高温胁迫对落叶果树需热量的影响研究较少[12]。【本研究切入点】本试验以不进行高温水浴处理作为对照,分析40、50、60、70 ℃高温水浴下5个品种葡萄休眠芽的萌发进程(膨大、露绿、展叶)和需热量的变化,以萌发进程和需热量共同来确定高温对葡萄芽休眠的打破情况。【拟解决的关键问题】旨在明确不同高温水浴热胁迫对葡萄芽休眠及其萌发的影响,为人工促进葡萄芽的休眠结束和萌发提供理论基础。
1 材料与方法
1.1 试验材料与试验设计
以4年生红地球、美人指、圣诞玫瑰、奥古斯特、玫瑰香葡萄枝条(自然休眠状态)为试验材料,于2016年11月20日(此时低温积累量约为需冷量的1/2)从宁夏大学试验基地贺兰园艺产业园15号阴阳棚采集。选择健壮的一年生枝条,每个品种取30根,每根枝条上有3~5个饱满的冬芽。采回到实验室后,用清水将葡萄枝条清洗干净,以试管架固定枝条,插在清水中,放入人工气候箱培养(SPX-250D型),温度25/15 ℃(昼夜),每5 d换1次水。11月20日到11月30日进行40、50、60、70 ℃的枝条底部0~10 cm水浴加热,每天1次,持续时间为1 h/次,以枝条底部0~10 cm不进行水浴加热为对照,每个处理组6根枝条,水域加热处理10次。2017年1月8日开始进行枝条芽的萌发观测。

表1 试验材料
1.2 试验方法
1.2.1 萌芽进度调查 参考Hellman[13]的结果,将葡萄枝条芽的萌发过程分为3阶段。膨大期:从芽膨大变圆、明显有别于初始的芽开始,到芽被绒毛覆盖,看不到嫩叶的颜色。统计每根枝条上处于膨大期及以后阶段的芽数目,膨大率(%)=(≥膨大期芽的数目)/总芽数×100。露绿期:从新芽的颜色能透过绒毛看到,到芽最外面一片叶子的边缘可见。统计每根枝条上处于露绿期及以后阶段芽的数目,露绿率(%)=(≥露绿期芽的数目)/总芽数×100。展叶期:从新芽最外面一片叶子的边缘可见开始,到第一片叶子完全展开。统计每根枝条上处于展叶期及以后阶段的芽数目,展叶率(%)=(≥展叶期芽的数目)/总芽数×100。
1.2.2 需热量 葡萄从自然休眠结束至50 %芽萌发展叶所需的热量累积,常用生长度时模型、有效积温模型计算。生长度时模型:该模型对需热量的估算用生长度小时(GDH ℃)表示:每1 h的温度所相当的热量单位(GDH ℃)根据下式计算。
GDH℃ = 0t≤4.5 ℃
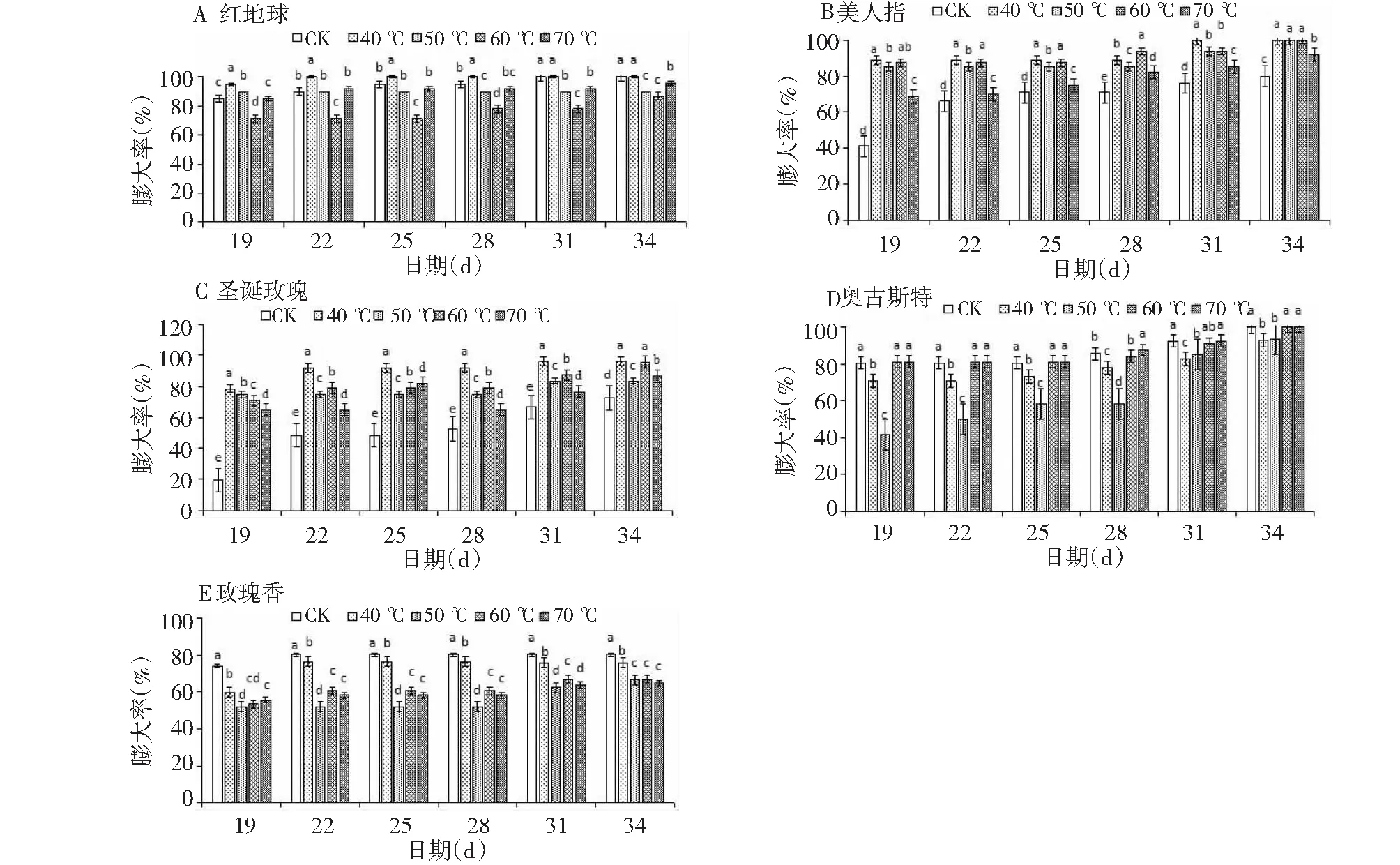
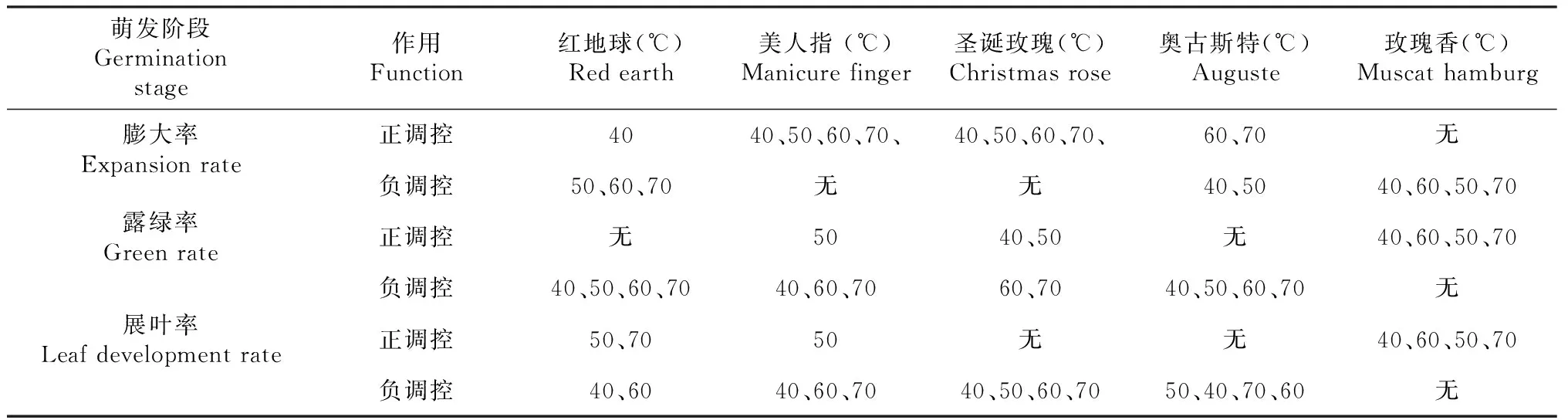
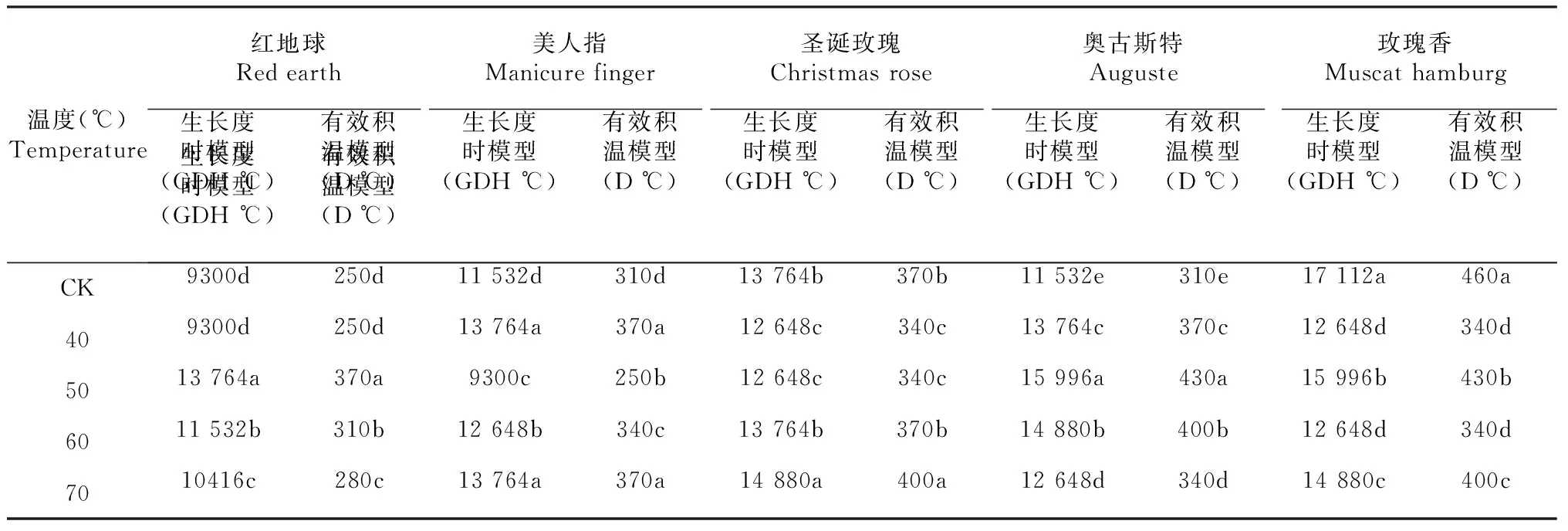
GDH℃ =t-4.5 4.5 ℃ GDH℃ = 20.5t≥25.0 ℃ 有效积温模型:该模型对需热量的估算用有效积温进行,单位为D ℃,它根据植物的生物学零度统计。根据下式计算:需热量(D ℃)=∑(日平均气温-生物学零度),葡萄的生物学零度为10 ℃。 1.2.3 数据分析 Excel 作图作表,DPS软件进行方差分析和拟合分析。 从图1可以看出,短时间高温处理对葡萄芽膨大率的影响因葡萄品种、温度的不同而异。40 ℃处理对红地球芽的膨大率具有正调控作用,其膨大率均显著性高于对照,而50、60、70 ℃处理则对于红地球芽的膨大率则表现出负调控作用,其膨大率均显著性低于对照(图1-A);40、50、60、70 ℃处理对美人指和圣诞玫瑰芽的膨大率具有正调控作用,其膨大率均显著高于对照,且40 ℃的效果最佳(图1-BC); 40、50 ℃处理对奥古斯特芽的膨大率具有负调控作用,二者膨大率均显著性低于对照,60、70 ℃处理对奥古斯特芽的膨大率具有正调控作用,二者的膨大率均显著性高于对照(图1-D);40、50、60、70 ℃处理对玫瑰香芽的膨大率具有负调控作用,其膨大率均显著性低于对照,且50 ℃的效果最好(图1-E)。 同一组中不同小写字母表示(P<0.05)差异显著图1 5个葡萄品种在不同高温下的膨大率变化Fig.1 Change of swelling rate of five grape varieties under different high temperature 从图2可以看出,短时间高温处理对葡萄芽露绿率的影响因葡萄品种、温度不同而异。40、50、60、70 ℃处理则对于红地球芽的露绿率均表现出负调控作用,其膨大率均显著性低于对照(图2-A);50 ℃处理对美人指芽的露绿率具有正调控作用,其露绿率均显著高于对照,40、60、70 ℃处理则对于美人指芽的露绿率具有负调控作用,其露绿率均显著性低于对照(图2-B); 40、50 ℃处理对圣诞玫瑰芽的露绿率具有正调控作用,其露绿率均显著高于对照,50 ℃的效果更好,60、70 ℃处理则对于圣诞玫瑰枝条芽的露绿率表现出负调控作用(图2-C);40、50、60、70 ℃处理对奥古斯特芽的露绿率具有负调控作用,其露绿率均显著性低于对照(图2-D);40、50、60、70 ℃处理对玫瑰香芽的露绿率具有正调控作用,其露绿率均显著性高于对照,且50 ℃的效果最好(图2-E)。 从图3可以看出,短时间高温处理对葡萄芽展叶率的影响因葡萄品种、温度不同而异。50、70 ℃处理则对于红地球芽的露绿率均表现出正调控作用,其展叶率均显著性高于对照,且70 ℃效果更好,40、60 ℃处理则对红地球芽的展叶率表现出负调控作用,其展展叶率均显著性低于对照(图3-A);50 ℃处理对美人指芽的展叶率有正调控作用,其展叶率均显著高于对照,40、60、70 ℃处理则对于美人指芽的展叶率均表现出负调控作用,其展叶率均显著性低于对照(图3-B);40、50、60、70 ℃处理则对于圣诞玫瑰芽的展叶率具有负调控作用,它们的展叶率均显著性低于对照(图3-C);40、50、60、70 ℃处理对奥古斯特芽的展叶率具有负调控作用,其展叶率均显著性低于对照(图3-D);40、50、60、70 ℃处理对玫瑰香芽的展叶率具有正调控作用,其展叶率均显著性高于对照,且50 ℃的效果最好(图3-E)。 同一组中不同小写字母表示(P<0.05)差异显著图2 5个葡萄品种在不同高温下的露绿率变化Fig.2 Changes of green rate of 5 grape varieties under different high temperature 同一组中不同小写字母表示(P<0.05)差异显著图3 5个葡萄品种在高温下的展叶率变化Fig.3 Change of leaf spreading rate of 5 grape varieties under different high temperature 萌发阶段Germination stage作用Function红地球(℃) Red earth美人指 (℃) Manicure finger圣诞玫瑰(℃) Christmas rose奥古斯特(℃) Auguste玫瑰香(℃) Muscat hamburg膨大率Expansion rate正调控4040、50、60、70、40、50、60、70、60、70无负调控50、60、70无无40、5040、60、50、70露绿率Green rate正调控无5040、50无40、60、50、70负调控40、50、60、7040、60、7060、7040、50、60、70无展叶率Leaf development rate正调控50、7050无无40、60、50、70负调控40、6040、60、7040、50、60、7050、40、70、60无 从表2可以看出,短时间不同高温处理对不同品种葡萄芽的膨大率、露绿率、展叶率的进程。相同高温处理对同一葡萄品种芽膨大率、露绿率、展叶率的调控效应不相同,如40 ℃对红地球枝条芽的膨大率、展叶率具有正调控效应,但其对红地球枝条芽的露绿率却表现出负调控效应。相同高温处理对不同葡萄品种芽的膨大率、露绿率、展叶率的调控效应相同,如40 ℃对红地球芽的膨大率、展叶率具有正调控效应,但40 ℃对奥古斯特芽的膨大率、展叶率却表现出负调控效应。不同高温处理对不同葡萄品种芽自然休眠解除的调控效应不同,所以造成了同一高温处理下,不同品种葡萄芽的萌发情况不同;相同温度对同一品种葡萄芽的膨大率、露绿率、展叶率的调控效应不同,可能是因为其生理机制的差异。 从表3可以看出,短时间高温处理对葡萄需热量的影响因品种、温度的不同而异,且2种模型统计的需热量结果趋势相同。不同高温对同一品种葡萄需热量的影响不同,如对于红地球、美人指和奥古斯特葡萄,高温处理对于其芽需热量具有正调控效应,即增加其需热量;对于圣诞玫瑰葡萄,40、50 ℃处理对于其芽需热量具有负调控效应,即减少其需热量;对于玫瑰香葡萄高温处理对于其芽需热量均具有负调控效应,即减少其需热量。同一高温对不同品种葡萄需热量的影响不同,如40 ℃处理对美人指的需热量具有正调控效应,对圣诞玫瑰、奥古斯特、玫瑰香葡萄却表现出负调控效应。 对葡萄休眠芽的膨大率有促进作用的高温处理不同。40 ℃处理对红地球休眠芽的膨大率具有正调控作用;40、50、60、70 ℃处理对美人指和圣诞玫瑰芽的膨大率具有正调控作用,且40 ℃的效果最佳;60、70 ℃处理对奥古斯特芽的膨大率具有正调控作用;40、50、60、70 ℃处理对玫瑰香芽的膨大率均无正调控作用,且50 ℃的效果最好。 对葡萄休眠芽露绿率有促进作用的高温处理不同。40、50、60、70 ℃处理则对于红地球芽的露绿率均无正调控作用;50 ℃处理对美人指芽的露绿率具有正调控作用; 40、50 ℃处理对圣诞玫瑰芽的露绿率具有正调控作用,50 ℃的效果更好;40、50、60、70 ℃处理对奥古斯特芽的露绿率无正调控作用;40、50、60、70 ℃处理对玫瑰香芽的露绿率具有正调控作用,且50 ℃的效果最好。 表3 不同高温处理下枝条芽的需热量 注:同一列中不同小写字母表示(P<0.05)差异显著。 Notes: Lowercase letters in same column indicate the significance at 0.05 level. 对葡萄休眠芽的展叶率有促进作用的高温处理不同。50、70 ℃处理则对于红地球芽的露绿率均表现出正调控作用, 70 ℃效果更好;50 ℃处理对美人指芽的展叶率有正调控作用;40、50、60、70 ℃处理则对于圣诞玫瑰芽的展叶率无正调控作用;40、50、60、70 ℃处理对奥古斯特芽的展叶率无正调控作用;40、50、60、70 ℃处理对玫瑰香芽的展叶率具有正调控作用,且50 ℃的效果最好。 短时间范围内一定的高温处理对葡萄芽需热量有影响,且有效积温模型和生长度时模型统计的需热量结果趋势相同。对于红地球、美人指和奥古斯特葡萄,高温处理对于其需热量具有正调控效应,即增加其需热量;对于圣诞玫瑰葡萄,40、50 ℃处理对于其芽需热量具有负调控效应,即减少其需热量;对于玫瑰香葡萄高温处理对于其芽需热量均具有负调控效应,即减少其需热量。 落叶果树的自然休眠是指在适宜的环境条件其芽仍不能萌发展叶,即使萌发展叶也不整齐[3]。1996年,Burr[14]用50~55 ℃对葡萄的休眠芽进行不同时间的水浴加热处理,发现50 ℃处理60~90 min对芽的自然休眠具有打破作用,55 ℃处理30 min以上时则对葡萄芽的萌发展叶表现出副作用并会伤害芽。Wample[7]对赤霞珠葡萄休眠枝条进行水浴加热处理,发现52~60 ℃对芽的萌发表现出负调控作用。王海波[8]以桃休眠桃枝条作为试验材料,发现高温能打破芽休眠并促进其萌发展叶。Halay[15]以Early sweet葡萄和Sugarone葡萄作为试验材料,发现以空气作为介质较以水作为介质进行50 ℃高温处理时对芽休眠解除的效果更好,还发现在150 ℃的半致死温度下,10~30 s就可以打破芽的自然休眠。但以上学者的研究材料较单一,对高温与芽萌发关系的研究有局限性。 本试验采用5个葡萄品种,对它们的葡萄底部0~10 cm进行水浴高温处理。试验结果表明,短时间的部分高温处理能够解除部分品种葡萄芽的自然休眠,进而促进葡萄芽的萌发展叶,但部分高温处理对部分品种葡萄的自然休眠解除没有效果,甚至伤害芽而延迟葡萄芽的萌发展叶(表2)。 葡萄的需热量是其萌芽展叶50 %所需的热量累积,用它来评估高温处理对葡萄芽的自然休眠解除效果时:40、50、60、70 ℃高温处理对红地球、美人指、奥古斯特葡萄的休眠解除没有效果,不能减少葡萄的需热量;40、50 ℃对圣诞玫瑰葡萄的休眠解除有效果,能减少葡萄的需热量;40、50、60、70 ℃高温处理对玫瑰香葡萄的休眠解除有效果,能减少葡萄的需热量。即相同高温处理对葡萄芽的休眠解除效果会随着品种的变化而发生变化。 自然条件下,冬季温度达到50 ℃几乎不可能,但在现代化设施内,通过加温措施可以达到,这对葡萄的栽培和繁殖具有重要的意义。2 结果与分析
2.1 水浴热胁迫对枝条芽膨大的影响
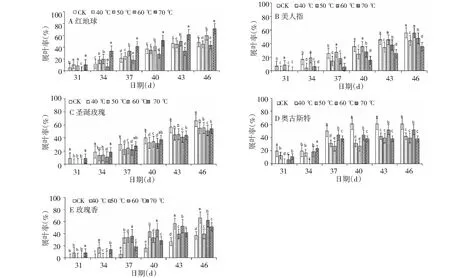
2.2 水浴热胁迫对枝条芽露绿的影响
2.3 水浴热胁迫对枝条芽展叶的影响

2.4 水浴热胁迫对枝条芽萌发进程的对比分析
2.5 水浴热胁迫对枝条芽需热量的影响
3 小 结
4 讨 论
猜你喜欢
今日农业(2022年16期)2022-11-09
环球时报(2022-05-26)2022-05-26
中学生数理化·中考版(2021年9期)2021-11-20
军事文摘(2021年16期)2021-11-05
小学科学(学生版)(2020年10期)2020-10-28
童话世界(2019年31期)2019-11-25
中学生数理化·中考版(2019年9期)2019-11-25
中华诗词(2019年12期)2019-09-21
小学生必读(低年级版)(2019年5期)2019-08-30
中华诗词(2019年1期)2019-08-23
