
蔬菜花青苷生物合成及转录调控的研究进展
2019-01-13 09:51曹琳娇李晓杰焦棒棒梁毅马长生
中国瓜菜 2019年12期
关键词:蔬菜
曹琳娇 李晓杰 焦棒棒 梁毅 马长生

摘 要:蔬菜在人们日常饮食中占据重要地位,可为人体提供维生素、礦物质等多种营养物质。颜色是蔬菜育种中重要的感官指标,而蔬菜颜色的形成受不同类型花青苷的影响。花青苷在植物生长发育、延缓机体衰老、预防心脏病等方面都发挥重要作用。研究表明,花青苷的生物合成受结构基因和调节基因的共同调控,分子水平上对蔬菜花青苷遗传机制的研究也在不断加深。笔者综述了与蔬菜花青苷生物合成途径中相关结构基因及其转录调控基因的研究现状,并对蔬菜花青苷的研究前景进行了展望。
关键词:蔬菜;花青苷;结构基因;转录调控
Advances in biosynthesis and transcriptional regulation of anthocyanin in vegetables
CAO Linjiao1, LI Xiaojie2, JIAO Bangbang1, LIANG Yi2, MA Changsheng1
(1. College of Horticulture, Henan Agricultural University, Zhengzhou 450002, Henan, China; 2. Beijing Vegetable Research Center, Beijing Academy of Agriculture and Forestry, Beijing 100097, China)
Abstract: Vegetables occupy an important position in people's daily diet, and its can provide a variety of nutrition material such as vitamins, minerals for human body. Color is an important sensory index in vegetable breeding, and the formation of vegetable color is influenced by different types of anthocyanins. Anthocyanin plays an important role in plant growth and development, delaying aging, preventing heart disease and other aspects of human health. Studies have shown that the biosynthesis of anthocyanin is co-regulated by structural genes and regulatory genes, and the research on the genetic mechanism of anthocyanin in vegetables at the molecular level is deepening. In this paper, the research status of structural genes and transcriptional regulation genes related to vegetable anthocyanin biosynthesis pathway is reviewed, and the research prospect of vegetable anthocyanin is prospected.
Key words: Vegetable; Anthocyanin ;Structural genes; Transcriptional regulatory
蔬菜是一年或多年生草本或木本植物,含有幼嫩多汁的食用器官。从植物学分类上看,我国普遍栽培的蔬菜分为十字花科、伞形科、茄科等20多个科,包括白菜、胡萝卜、番茄等物种。蔬菜中含有大量的花青素、类胡萝卜素、番茄红素、维生素C等营养元素,具有较高的食用价值,还有增加饱腹感、促进消化等作用,是人体吸收膳食纤维、矿物质等营养物质的主要来源。其中,深色蔬菜(深绿色、红色、橘红色、紫红色等)更具营养优势。这些深色蔬菜由于高度富含花青素而越来越受到消费者的青睐,蔬菜颜色也成为选育和食用蔬菜最重要的品质之一。
花青素是植物次生代谢产物,属于类黄酮物质,主要积累在叶片、果实和花瓣表皮细胞的液泡内,赋予植物蓝、紫、红等颜色。花青素主要分为天竺葵素、矢车菊素、飞燕草素、芍药花素、矮牵牛素和锦葵色素[1],如紫白菜中高度酰基化和糖基化修饰的矢车菊素[2],洋葱中的矢车菊色素和飞燕草色素以及心里美萝卜中的天竺葵素[3]。但花青素在植物内不能稳定存在,常与糖(葡萄糖、半乳糖、芸香糖、龙胆三糖等)通过糖苷键形成花青苷。
花青苷在调控生长素运输、花的发育等方面发挥作用。花青苷生物合成过程中产生的黄酮醇已被认为与生长素转运负调控有关;花青苷还可能通过增加生长素的细胞内浓度,转而促进植物极性生长。花青苷能使植物的花呈现五颜六色,有利于吸引昆虫传播花粉,也有利于鸟类和动物采食种子以促进种子的传播和繁育。花青苷还广泛参与了防御紫外线、植物与微生物的互作、植物抗逆等过程。花青苷具有多种生物活性功能和药用价值,因其有较强的氧化还原活性,可作为抗氧化剂、自由基清除剂和二价阳离子的螯合剂,用于延缓人类机体衰老。此外,还有研究发现,花青苷在保护视力、预防心脏病及降低癌症等方面发挥非常重要的医疗保健功能。
花青苷的生物合成受两类基因的共同控制,一类是结构基因,编码其生物合成途径中所需要的酶;另一类是调节基因,其编码的转录因子调控结构基因的时空表达[4]。其生物合成途径是植物中表征较为完善的次级代谢途径之一,合成途径中所包含的主要结构基因和许多调控基因已经在拟南芥、玉米、矮牵牛等模式作物中得到了很好的表征。目前对蔬菜花青苷的研究不仅落后于模式作物,也落后于果树、花卉等园艺作物,但关于蔬菜花青苷的积累特性、生物合成及调控机制也取得了一定的进展。笔者概述了蔬菜花青苷生物合成途径关键结构基因及其转录调控基因的研究现状。
1 与花青苷合成相关的结构基因
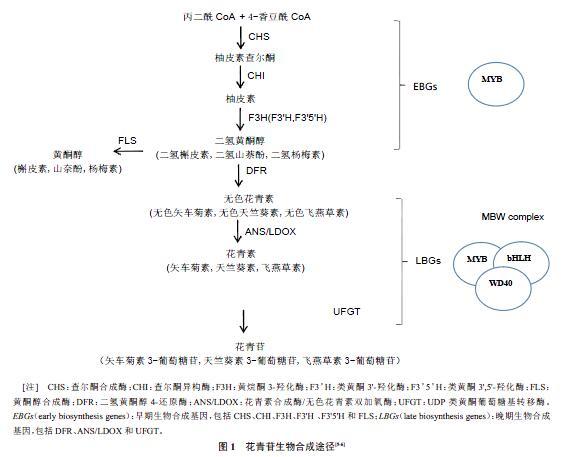
花青苷的生物合成是一系列酶促反应(图1),其生物合成途径已经在马铃薯、番茄、茄子、洋葱、甘蓝等蔬菜作物中都有了一定的研究。
1.1 CHS(Chalcone synthase)
CHS是类黄酮代谢途径中的第一个关键限速酶,位于花青苷合成通路的上游,催化4-香豆酰辅酶A和对香豆酰CoA生成查尔酮。甘薯IbCHS1[7]、茄子SmCHS[8]、辣椒CaCHS04和CaCHS05[9]等基因的表达量与不同颜色品种中花青苷含量呈正相关,表明这些CHS基因在花青苷合成途径中起关键作用。然而,胡朝阳等[10]通过RT-PCR分析表明,紫色马铃薯StCHS基因主要在茎和叶柄中表达,而在根、块茎和叶轴中几乎检测不到,表明StCHS基因表达具有组织特异性。
1.2 CHI(Chalcone isomerase)
查尔酮经CHI催化生成二氢黄酮醇,CHI基因表达量的高低通过影响查尔酮代谢的量间接影响花青苷的合成。CHI基因一般在植物各个组织中都有表达,其中白菜CHI(BrCHI)基因家族的表达还存在一定的器官差异性和时空特异性。如BrCHI1在生殖器官中表达;而BrCHI2主要在发育早期的种子中表达;BrCHI3在叶、花、蕾和中期种子中表达;而BrCHI4 也是主要在生殖器官中表达,但在种子发育后期表达量明显高于其他阶段。BrCHI基因各成员间表达的时空特异性表现在:BrCHIl表达量最高,BrCHI4表达量最低。洋葱CHI基因的失活阻断了花青苷的生物合成,导致了金色鳞茎的产生,其在番茄中的共表达可以增强花青素和黄酮醇的产生[11],而且它对拟南芥PAP1调控基因的协同作用可以增强番茄皮的花青苷含量[12]。Guo J等[13]的研究结果表明,IbCHI是甘薯花青苷生物合成途径中发挥作用的关键酶,在根发育的早期阶段负责花青苷生物合成的活化。
1.3 F3H(Flavanone 3-hydroxylase)
F3H位于花青苷合成途径的中枢位点,控制着合成途径的代谢流向,在种皮和花的颜色形成中具有重要作用。F3H基因在不同的植物或组织中具有底物特异性,可以催化黄烷酮在C3位置羟基化形成无色的黄烷酮醇,也能够催化圣草酚、柚皮素、5羟基双氢黄酮等在3位置的羟基化生成二氢黄酮醇。F3H基因在大多数物种中也仅以单拷贝形式存在,但在洋葱中以双拷贝形式存在,并催化柚皮素羟基化以产生二氢山萘酚。BrcF3H在紫色不结球白菜叶片中的表达量极显著高于在绿色叶片中的表达量,且其表达水平与不结球白菜叶片紫色着色程度正相关,推测此F3H基因为影响不结球白菜花青苷生物合成的关键基因之一。光在F3H基因的表达中发挥重要作用,如光可以诱導芜菁F3H基因的表达;闫瑞霞等[14]也通过叶片遮光处理试验证明紫山药DaF3H基因受光调节,且DaF3H基因表达与花青苷的积累表现为同步。
1.4 DFR(Dihydroflavonol 4-reductase)
DFR是花青苷合成后期发挥关键作用的酶,是决定无色到有色的重要调控点,DFR基因在进化中高度保守。Sunggil kim等[15-16]利用RACE技术克隆了DFR基因,通过RT-PCR分析发现DFR基因仅在红皮洋葱中表达,转录水平上DFR基因的失活导致美国黄洋葱中缺乏花青苷,并进一步根据DFR基因的突变设计了基于PCR的分子标记。Sunggil kim等[17]研究发现了DFR-A基因的2个新的突变类型也会导致洋葱表皮花青苷的缺失;Song等[18]研究发现,至少有9个DFR-A基因独立的自然突变会导致黄皮洋葱中花青苷合成的缺失。Zhang Chunsha等[19]通过转录组测序和代谢分析表明,DFR基因在洋葱深红色鳞茎的生物合成中发挥重要作用,其表达水平可能起到阻止蓝色色素沉着的作用。郭晋雅等[20]研究发现,紫肉甘薯IbDFR在块根中的表达量与其花青苷含量的变化趋势一致;Hongxia W等[21]用RNAi方法下调了转基因甘薯中IbDFR基因的表达,则显著减少了甘薯幼叶、茎和贮藏根中花青苷的积累。这都表明IbDFR可能是紫甘薯花青苷合成代谢过程中发挥关键作用的酶基因。SmDFR基因、BrcDFR基因也分别在茄子和不结球白菜花青苷生物合成中发挥重要作用。
1.5 ANS(Anthocyanidin synthase)
ANS是位于花青苷合成通路末端的酶,催化无色花青素向有色花青素的转化,其在基因水平、转录水平、蛋白水平存在的差异都有可能会影响花青苷的合成。Eun-Young Kim等[22]鉴定出2个新的ANS基因的等位变异,这种变异可以造成洋葱花青苷合成的缺失。许玉超等[23]发现,紫色不结球白菜ANS基因(BrcANS)mRNA表达量与叶片紫色直接相关,表明其可能在转录水平上调控叶片中紫色的形成。孙玉燕等[24]的研究表明,心里美萝卜RsANS推导得到的氨基酸序列与白萝卜存在4个氨基酸的差异,这种差异可能是导致心里美萝卜和白萝卜根肉色不同的关键因素之一。
1.6 UFGT(UDP-flavonoid glucosyltransferase)
UFGT是花青苷合成途径发挥作用的最后一个关键限速酶。张洪伟等[25]从洋葱中克隆得到了AcUFGT1基因和AcUFGT2基因,表达分析的结果表明,这2个基因在红洋葱中表达量很高,但在黄洋葱和白洋葱中几乎不表达,这表明AcUFGT1和AcUFGT2与洋葱鳞茎不同颜色的形成有关。前人通过紫肉甘薯和黄肉甘薯的RNA测序结果对比、PCR检测和Southern blot等方法发现并证明了UF3GT是甘薯花青苷合成途径的关键酶之一,且3GT是紫肉甘薯花青苷的重要组分之一。Liu Y等[26]证明了StUFGT1与马铃薯块茎中的花青苷生物合成相关,还发现了11个注释为UFGT的基因在紫色薯块与薯皮中差异表达;RNA-seq结果表明,紫色和白色马铃薯品种的UFGT基因有涉及功能变化或基因定位的SNP。
2 与花青苷合成相关的调节基因
基因的时空表达受转录水平、转录后水平和翻译水平的调控,而类黄酮生物合成的调节点通常发生在转录水平。参与花青苷生物合成的主要调节基因如图2所示。
2.1 R2R3-MYB转录因子
MYB蛋白是高度保守的,所以调控基因的特异性较高,该结构域通常包括1~4个氨基酸重复(R基序)。MYB蛋白每个R基序中第3个α螺旋在与DNA接触过程中起到识别DNA的功能,该α螺旋可以结合到DNA双螺旋的大沟结构内,能使MYB蛋白精确的结合到特定DNA序列上。根据R基序的不同,可以將MYB分为R2R3-MYB、1R-MYB、3R-MYB和4R-MYB这4类。其中,编码R2R3-MYB转录因子的基因被认为是决定植物中黄酮类化合物空间和时间发生的关键。现已从许多蔬菜中分离并鉴定了参与调控花青苷合成的R2R3-MYB转录因子,如洋葱AcMYB1、白菜BrMYB2和BrMYB4、甘薯IbMYB1等。
Schwinn Kathy E等[29]证明了AcMYB1是洋葱产生花青苷的正向调节因子,是洋葱鳞茎产生红色表型的关键;短暂过表达AcMYB1基因时可诱导洋葱组织中花青苷的产生,且通过RNAi瞬时抑制其表达时可以减少色素沉着。穆春等[30]的研究表明,白菜BrMYB2和BrMYB4及甘薯IbMYB1也在花青苷积累中发挥正调控作用,其中BrMYB2还参与白菜花青苷生物合成的激活过程;IbMYB1被认为可能参与调控了IbCHI、IbDFR等与花青苷合成相关的结构基因。Chu H等[3]随后发现,甘薯IbMYB1a基因的表达可以诱导拟南芥中的花青素积累,且其可以诱导ANS基因表达上调。大部分R2R3-MYB转录因子对结构基因的表达起正调控作用,还有少数起负调控作用。如草莓FaMYB1(R2R3-MYB)在烟草中的异源表达就会抑制烟草花瓣中花青素及槲皮素的积累,并影响ANS、UFGT基因的表达活性[32]。
2.2 bHLH(basic helix-loop-helix)
bHLH是根据其蛋白保守结构域命名的一类转录因子家族,在植物中广泛存在,通过调控花青苷合成途径的一些结构基因的表达,使花青苷在植物细胞中积累[33]。在bHLH结构域的碱性氨基酸序列中,HER(His5-Glu9-Arg13)基序具有高度的保守性,可结合启动子序列中的六核苷酸E-box基序,进而调控靶基因转录。白菜BrTT8的表达量与花青苷积累程度保持一致,如在红菜薹花青苷高度积累的组织中表达量较高,在无花青苷积累的红菜薹组织和少花青苷积累的小白菜中的表达量较低。BrTI8的表达模式与ANS基因在红菜薹花青苷合成过程中的表达模式相一致,表明BrTI8可能参与调控ANS基因的表达。
有些bHLH也对花青苷积累起抑制作用。如玉米INI(其编码的蛋白与玉米bHLH部分同源)缺失突变体中,UFGT基因表达量和花青苷含量明显提高。杨琳等[34]的研究表明,bHLH可能通过与bHLH蛋白形成异源二聚体或与无活性的MYB蛋白结合,又或者与转录复合物的下游靶基因直接结合干扰活性bHLH-MYB的形成或阻碍MYB复合物发挥激活功能而抑制花青苷合成。
2.3 WD40
WD40是一类含4~10个随机WD重复结构域的蛋白质家族,该结构域由总长约40个以色氨酸(W)和天冬氨酸(D)结尾的氨基酸序列组成。Wei Dong等[35]从甘薯分离得到的IbWD40基因的表达与不同甘薯品种的花青素积累呈正相关;该基因在转基因拟南芥幼苗中表达上调并积累了花青苷,表明IbWD40在紫色肉质甘薯花青苷生物合成的调节中起关键作用。Yongfeng G等[36]发现,番茄SlAN11基因过表达时会抑制SlFLS的表达从而减少花青苷合成途径中间产物黄酮醇或其糖基化衍生物的积累;还发现了SlAN11与bHLH蛋白互作(SlAN11不与MYB蛋白互作,而bHLH与MYB互作),并对番茄黄酮类化合物的合成和种子休眠具有重要的调控作用。
2.4 MBW
MBW(MYB-bHLH-WD40)转录复合物通常由R2R3-MYB、bHLH和WD40共同组成。R2R3-MYB通常需要与bHLH和WD40蛋白相互作用来充分发挥其转录调控花青苷合成的功能,其R3内的[DE]Lx2[RK]x3Lx6Lx3R基序能与bHLH 结合并互作;bHLH中也有与MYB结合的重要结构域;而WD40则起着增强MBW复合体稳定性的作用。王冠杰[37]通过酵母双杂交试验表明,芜菁BrMYB75与BrTT8互作,并通过瞬时转染烟草叶片表明BrMYB75与BrTT8协同调控芜菁CHS基因的表达。Wenjun Huang等[38]的研究发现并证明了EsMYB1可以与几个bHLH调控因子互作并激活DFR和ANS基因的表达,通过上调DFR和ANS基因的表达来实现对花青苷合成的调控。Yanjie Zhang等[39]的试验表明,区域特异性组装的功能性MBW(由BrTT8、SlAN2和SlAN11组成)蛋白复合物使Pro35S:BrTT8番茄果实在强光诱导下显示不均匀的花青苷色素沉着。
2.5 其他调控因子
花青苷生物合成途径会受到R3-MYB转录因子和N / NO(3)(-)诱导的LBD(lateral organ boundary domain)转录因子家族的负调控。R3-MYB通过扰乱MBW复合物形成而抑制花青苷合成;Albert N W等[40]通过酵母三杂交试验也证实了MYB激活因子和抑制因子可同时与相同的bHLH结合。拟南芥MBW转录激活物的复合物激活拟南芥同源域蛋白基因GLABRA2(GL2),促进毛状体形成和抑制根毛形成;此MBW复合物还可以激活R3-MYB基因,被激活的R3-MYB通过与R2R3-MYB蛋白竞争结合bHLH蛋白而发挥负反馈作用,阻断MBW复合物的形成[41]。LBD转录因子家族包括LBD37、LBD38和LBD39基因;当没有N / NO(3)(-)存在时,3种LBD基因中的任意一个过表达都会强烈抑制拟南芥花青苷合成基因PAP1和PAP2的表达[42]。董慧杰等[43]研究表明,不结球白菜中花青苷负调控基因BrcLBD39与叶片中花青苷的积累密切相关,在花青苷的生物合成过程中起着重要的调控作用。
3 展 望
关于茄子、白菜、洋葱、马铃薯等蔬菜花青苷生物合成及转录调控的研究已经取得了一些进展,克隆和鉴定出很多和花青苷合成相关的结构基因和调节基因,但仍有很大的发展空间。与模式作物相比,参与蔬菜花青苷合成的结构基因和调节基因可能更具复杂性和多变性。
且高通量测序技术、蛋白质组学和代谢组学等多组学分析技术的发展也为蔬菜花青苷的研究提供了更多的思路和方法。此外,光、温度等环境因素也影响了花青苷的形成。蔬菜花青苷的研究应当紧密联系实际,为分子改良富含高花青苷的新品种和改良蔬菜颜色等夯实遗传基础;并通過分子设计育种的手段加速育种进程,培育高品质、有特色的新品种,为研究相关蔬菜作物的花青苷合成分子机制提供重要的理论价值。总而言之,蔬菜花青苷生物合成途径及其调控机制还需要不断地探索和研究,以便充分发挥花青苷的食用功能,创造更大的价值。
参考文献
[1] 陈敏,杨清,CHENMIN,等.马铃薯花色苷生物合成与调控研究进展[J].中国马铃薯,2013,27(4):232-238.
[2] 张彦杰.紫白菜与紫茄花色苷生物合成及光调控的分子机理研究[D].重庆:重庆大学,2015.
[3] 穆春,王丽,贾晓琳,等.RNAi抑制过氧化物酶基因Rsprx1表达促进萝卜花青素的积累[J].中国生物化学与分子生物学报,2013,29(9):867-872.
[4] GROTEWOLD E.The genetics and biochemistry of floral pigments[J].Annu.Rev.Plant Biol,2006,57(1):761-780.
[5] PETRONI K,TONELLI C.Recent advances on the regulation of anthocyanin synthesis in reproductive organs[J].Plant Science,2011,181(3):219-229.
[6] 李益,张一卉,李化银,等.紫色芸薹属蔬菜花青素合成调控研究进展[J].山东农业科学,2014,46(11):137-142.
[7] 徐靖,朱家红,韩义胜,等.一个新的甘薯查尔酮异构酶基因的克隆和表达分析[J].基因组学与应用生物学,2018,37(2):845-849.
[8] ZHANG Y,HU Z,CHU G,et al.Anthocyanin Accumulation and Molecular Analysis of Anthocyanin Biosynthesis-Associated Genes in Eggplant (Solanum melongena L.)[J].Journal of Agricultural and Food Chemistry,2014,62(13):2906?2912.
[9] 徐婉莉,裴徐梨,荆赞革,等.辣椒查尔酮合成酶基因家族全基因组鉴定及表达特征分析[J].基因组学与应用生物学,2015,34(8):1747-1752.
[10] 胡朝阳,周友凤,龚一富,等.紫色马铃薯查尔酮合成酶基因(CHS)的克隆及分析[J].中国农业科学,2012,45(5):832-839.
[11] LIM W,LI J.Co-expression of onion chalcone isomerase in Del/Ros1-expressing tomato enhances anthocyanin and flavonol production[J].Plant Cell Tissue & Organ Culture,2016,128(1):1-12.
[12] LIM W,LI J.Synergetic effect of the Onion CHI gene on the PAP1 regulatory gene for enhancing the flavonoid profile of tomato skin[J].Scientific Reports,2017,7(1):12377.
[13] GUO J,ZHOU W,LU Z,et al.Isolation and Functional Analysis of Chalcone Isomerase Gene from Purple-Fleshed Sweet Potato[J].Plant Molecular Biology Reporter,2015,33(5):1451-1463.
[14] 闫瑞霞,殷剑美,韩晓勇,等.紫山药花青素调控基因DaF3H的克隆及表达分析[J].园艺学报,2014,41(4):701-712.
[15] KIM S,BINZEL M L,PARK S,et al.Inactivation of DFR (Dihydroflavonol 4-reductase) gene transcription results in blockage of anthocyanin production in yellow onions (Allium cepa).Molecular Breeding,2004,14(3):253-263.
[16] KIM S,YOOK S,PIKE L M.Development of a PCR-based marker utilizing a deletion mutation in the dihydroflavonol 4-reductase (DFR) gene responsible for the lack of anthocyanin production in yellow onions (Allium cepa)[J].Theoretical and Applied Genetics,2005,110(3):588-595.
[17] KIM S,BAEK D,CHO DY,et al.Identification of two novel inactive DFR-A alleles responsible for failure to produce anthocyanin and development of a simple PCR-based molecular marker for bulb color selection in onion (Allium cepa L.)[J].Theoretical and Applied Genetics,2009,118(7):1391-1399.
[18] SONG S,KIM C W,MOON J S,et al.At least nine independent natural mutations of the DFR - A gene are responsible for appearance of yellow onions ( Allium cepa L.) from red progenitors[J]. Molecular Breeding,2014,33(1):173-186.
[19] ZHANG C,LI X,ZHAN Z,et al.Transcriptome sequencing and metabolism analysis reveals the role of cyanidin metabolism in dark-red onion (Allium cepa L.) Bulbs[J].Scientific Reports,2018,8(1):14109.
[20] 郭晉雅,李云萍,傅玉凡,等.紫心甘薯二氢黄酮醇4-还原酶基因表达及酶活性与花色苷积累的相关性[J].中国农业科学,2011,44(8):1736-1744.
[21] WANG H,FAN W,LI H,et al.Functional characterization of dihydroflavonol-4-reductase in anthocyanin biosynthesis of purple sweet potato underlies the direct evidence of anthocyanins function against abiotic stresses[J].PLoS ONE,2013,8(11):e78484.
[22] KIM E Y,KIM C W,KIM S.Identification of two novel mutant ANS alleles responsible for inactivation of anthocyanidin synthase and failure of anthocyanin production in onion (Allium cepa L.)[J].Euphytica,2016,3(212):1573-5060.
[23] 许玉超,侯喜林,徐玮玮,等.紫色不结球白菜花色苷合酶基因BrcANS的克隆与表达分析[J].作物学报,2016,42(6): 850-859.
[24] 孙玉燕,段蒙蒙,邱杨,等.心里美萝卜花青素合成酶基因RsANS克隆及花青素生物合成相关基因表达分析[J].植物遗传资源学报,2016,17(5):889-896.
[25] 张洪伟,梁毅,刘小义,等.洋葱UFGT基因的克隆和表达分析[J].核农学报,2015,29(9):1677-1686.
[26] LIU Y,LIN-WANG K,DENG C,et al.Comparative Transcriptome Analysis of White and Purple Potato to Identify Genes Involved in Anthocyanin Biosynthesis[J].Plos One,2015,10(6):e0129148.
[27] XU W,DUBOS C,LEPINIEC L.Transcriptional control of flavonoid biosynthesis by MYB–bHLH–WDR complexess[J].Trends in Plant Science,2015,20(3):176-85.
[28] ZHENG T,TAN W,YANG H,et al.Regulation of anthocyanin accumulation via MYB75/HAT1/TPL-mediated transcriptional repressions[J].Plos One,2019,15(3):e1007993.
[29] SCHWINN KE,NGO H,KENEL F,et al.The onion (Allium cepa L.) R2R3-MYB gene MYB1 regulates anthocyanin biosynthesis[J].Frontiers in Plant Science,2016(7):1865.
[30] HE Q,ZHANG Z,ZHANG L.Anthocyanin Accumulation,Antioxidant Ability and Stability,and a Transcriptional Analysis of Anthocyanin Biosynthesis in Purple Heading Chinese Cabbage (Brassica rapa L.ssp.pekinensis )[J].Journal of Agricultural and Food Chemistry,2016,64(1):132-145.
[31] CHU H,JEONG JC,KIM WJ,et al.Expression of the sweetpotato R2R3-type IbMYB1a gene induces anthocyanin accumulation in Arabidopsis[J].Physiologia Plantarum,2013,148(2):189-199.
[32] MA D,CONSTABEL CP,et al.MYB Repressors as Regulators of Phenylpropanoid Metabolism in Plants[J].Trends in plant Science,2019,24(3):275-289.
[33] NUNO P,LIAM D.Origin and diversification of basic-helix-loop-helix proteins in plants[J].Molecular biology and evolution,2010,27(4):862-874.
[34] 杨琳,王宇,杨剑飞,等.花青素积累相关负调控因子的研究进展[J].园艺学报,2014,41(9):1873-1884.
[35] DONG W,NIU L,GU J,et al.Isolation of a WD40-repeat gene regulating anthocyanin biosynthesis in storage roots of purple-fleshed sweet potato[J].Acta Physiologiae Plantarum,2014,5(36):1861-1664.
[36] GAO Y,LUI J,CHEN Y,et al.Tomato SlAN11 regulates flavonoid biosynthesis and seed dormancy by interaction with bHLH proteins but not with MYB proteins[J].Horticulture Research,2018,5(1):1-18.
[37] 王冠杰.蕪菁MYB75和TT8在花青素合成途径中相互作用的分析[D].哈尔滨:东北林业大学,2013.
[38] HUANG W,SUN W,L? H,et al.A R2R3-MYB transcription factor from Epimedium sagittatum regulates the flavonoid biosynthetic pathway[J].PloS one,2013,8(8):e70778.
[39] ZHANG Y,LI Y,LI W,et al.Metabolic and molecular analysis of nonuniform anthocyanin pigmentation in tomato fruit under high light[J].Horticulture Research,2019,6:56.
[40] ALBERT N W,DAVIES K M,LEWIS D H,et al.A Conserved Network of Transcriptional Activators and Repressors Regulates Anthocyanin Pigmentation in Eudicots[J].Plant Cell,2014,26(9):962-80.
[41] ZHENG K,TIAN H,HU Q,et al.Ectopic expression of R3 MYB transcription factor gene OsTCL1 in Arabidopsis,but not rice,affects trichome and root hair formation[J].Scientific Reports,2016,6(1):19254.
[42] RUBIN G,TOHGE T,MATSUDA,et al.Members of the LBD family of transcription factors repress anthocyanin synthesis and affect additional nitrogen responses in Arabidopsis[J].Plant Cell,2009,21(11):3567-3584.
[43] 董慧杰,侯喜林,韩克,等.不结球白菜花青苷合成负调控基因BrcLBD39的克隆和表达分析及其对外源6-BA的响应[J].南京农业大学学报,2018,41(1):49-56.
猜你喜欢
小太阳画报(2020年3期)2020-04-24
中国总会计师(2017年1期)2017-03-10
广东第二课堂·小学(2017年2期)2017-02-20
广东第二课堂·小学(2017年1期)2017-02-20
儿童故事画报(2016年7期)2017-02-08
小学生·多元智能大王(2014年3期)2014-03-21
学苑创造·B版(2009年5期)2009-06-30
小雪花·小学生快乐作文(2009年4期)2009-04-27
