
探地雷达在典型高寒草地高原鼠兔洞道结构研究中的应用
2018-12-27 03:12宜树华
草业科学 2018年12期
秦 炎,秦 彧,宜树华,3
(1.中国科学院西北生态环境资源研究院,甘肃 兰州 730000; 2.中国科学院大学,北京 100049;3.南通大学,江苏 南通 226000)
高原鼠兔(Ochotonacurzoniae)是栖息在青藏高原的一种小型哺乳动物[1],通过采食、挖掘、排泄等活动参与草地生态系统能量流动和物质循环,其植食性特点和挖掘活动对高寒草地植被、生物多样性、物质循环等方面具有重要影响[2-3]。然而,目前对于高原鼠兔在高寒草地生态系统中的作用还存在巨大争议。一方面,高原鼠兔挖掘活动会破坏植被根系和土壤结构、加速土壤侵蚀、降低土壤肥力、减少植被生物量,增加与家畜在食物争夺中的竞争压力[4]。另一方面,有研究认为高原鼠兔的挖掘活动使得洞道外的生物及非生物物质通过洞口进入洞道,从而改变当地土壤结构和土壤通透性,使得原本较为封闭的土层与外界联通,较深层的水分、渗透率、通气性、容重及部分化学性质由此发生变化,其洞道在增加高原环境异质性等方面起着积极的作用[5]。因此,高原鼠兔也被认为是高寒草地生态系统的关键物种[6]。由于高原鼠兔营地下栖息,研究其洞道结构是认识高原鼠兔在高寒草地生态系统中作用的一个重要因素。鉴于此,有学者已经开展了鼠兔洞道结构的研究。例如,卫万荣等[7]通过对独立洞系的解剖,发现高原鼠兔洞道的特点是对高原环境的一种适应性选择;马波[8]针对高原鼠兔洞穴的特点采用舞台烟雾注入的方式,推断高原鼠兔洞穴结构的复杂度,研究高原鼠兔洞穴结构的空间分布特征。但目前就高原鼠兔洞道结构的研究比较匮乏,且前期研究需要实地挖取剖面,对生态环境具有一定的破坏性,加之高原鼠兔分布面积较广,加大了调查的难度。因此,寻求一种非破坏性且快速的洞道调查方法显得尤为重要。
探地雷达(ground penetrating radar,GPR)利用地面发射天线向地下发射短频脉冲,由接收天线接收回波信号,根据接收反射回波的双程走时、幅度相位等信息,对地下结构进行判别,求出目标深度[9],由于其无损探测的特性,被广泛应用在交通、石油勘探等领域。目前国内外学者在GPR的自动目标检测方面做了大量研究,庞希斌等[10]的研究显示了探地雷达在跑道检测中的快捷性、实用性、先进性。但在高寒草地小目标的应用研究还相对较少。就其对小目标探测的准确性和可行性,冯温雅等[11]指出探地雷达能够探测出浅地层中的小目标,为调查高原鼠兔洞道结构提供了可能。因此,本研究在实地调查不同样地类型高原鼠兔的洞道结构的基础上,分析不同样地类型高原鼠兔洞道结构的特点,并初步探讨探地雷达对探测高原鼠兔洞道结构的可行性,旨在量化高原鼠兔洞道结构特征,同时寻求一种无损、快速、大面积调查高原鼠兔洞道的新方法。
1 材料与方法
1.1 研究区概况
研究区位于青藏高原东北缘、祁连山西段,青海省海西蒙古族藏族自治州天峻县苏里乡,属疏勒河上游流域,流域面积1.1万km2,海拔2 078~5 763 m,其经纬度范围为38.2°-40.0° N,96.6°-99.0° E。该区属大陆性干旱荒漠气候区,气候干冷、多风,多年平均气温-2.7 ℃,年均降水量349.2 mm[12];当地植物种类主要有高山嵩草(Kobresiapygmaea)、苔草(Carextristachya)、铁棒锤(Aconitumpendulum)、紫菀(Astertataricus)、火绒草(Leontopodiumalpinum)、金露梅(Potentillafruticosa)、蒿草(Artemisavulgaris)和西伯利亚蓼(Polygonumsibiricum)等[13]。草地类型主要有沼泽草甸、高寒草甸、草原化草甸和高寒草原。由于多年冻土退化、放牧等综合因素的影响,研究区沼泽草甸和高寒草甸发生了不同程度的退化[14]。研究区域内高原鼠兔广泛分布,为研究其洞道结构及分布提供了便利的条件。
1.2 野外工作
1.2.1洞道结构调查 于2017年6-8月分别选择退化沼泽草甸(植被破碎化严重,多是孤立植被,裸地斑块已连通)、高寒草甸、退化草甸(植被破碎化严重,多是孤立植被,裸地斑块已连通)和高寒草原4种样地先后进行雷达探测和实地调查(图1)。在每种样地类型采用解剖方式的调查方法沿高原鼠兔洞道走向挖取4~5个土壤剖面,分别测定洞道的长度、深度、拐点、直径等结构特征,并结合实地剖析测量的数据,绘制洞系结构的俯视平面图并记录数据。根据记录的数据,绘制三维结构图。对不同植被类型下洞道结构差异进行分析,在此基础上,验证探地雷达的测量结果。

图1 疏勒河流域高原鼠兔洞道结构研究采样点Fig. 1 Sampling sites for plateau pika burrows at Shule River
1.2.2雷达探测 本研究采用中国电波传播研究所研发生产的探地雷达(LTD-2600),由主机系统和收发天线系统两大部分组成,收发天线系统采用的是400 MHz天线。通过实地调查发现鼠兔洞道的深度大多是在40 cm处,为了使雷达显示清晰且不丢失极个别特别深的鼠洞,增益大于0.4 m,又不至于信号接收器过载,调节增益至0.7 m,考虑到草地凹凸不平产生丢失接收电波信号的情况,采用测距轮的测量方法。此方法在拖动雷达时,每行走0.22 cm发射出一道电波信号,当遇到不同介质时一部分信号将会反射被主机接收,另一部分继续向下探测,如果接收到的信号数量少,主机会有速度过快的警告。对测距轮进行标定后,在高寒草甸(2个)、退化沼泽(1个)、高寒草原(3个)和退化高寒草甸(1个)的7个样地(图1)进行数据采集,分别随机选取3个10 m×10 m样方(合计21个,图2),在每个样方内以红色为起点,绿色为终点,采用来回测量方式(间隔为1 m),先进行横向的11次间断测量,再进行纵向的11次的测量。每个样方总共进行22次间断测量(图2),并记录样方内的洞口数量。

图2 探地雷达样方设置和测量方式 Fig. 2 Sample sites and measurement of GPR
GPR, ground penetating radar; similarly for the following figures.
1.2.3鼠兔洞口密度调查 在所设置的采样点,应用Yi[15]开发的无人机航拍系统对鼠兔密度进行调查,无人机航拍调查的步骤:1)新建工作点或选择已有工作点;2)根据采样点位置(图1)或选择已存在航线;3)设置无人机飞行高度(本研究中为20 m)、飞行速度(本研究中为4 m·s-1)及相机参数(白平衡,IOS等);4)上传航点、飞行高度与速度;5)无人机根据预设参数开始自主飞行与拍摄,对所有采样点进行航拍,每个采样点获取航拍照片16张,共获得112张照片[16]。每张照片覆盖面积约为26 m×35 m,每个像素的分辨率大约为1 cm,高原鼠兔的洞口的直径约为6 cm,调查过后,将数据整合到Mysql数据库中[3]。
1.3 数据分析
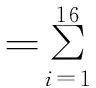

图3 探地雷达探测高原鼠兔洞道产生的波动图Fig. 3 GPR fluctuation of plateau pika tunnel
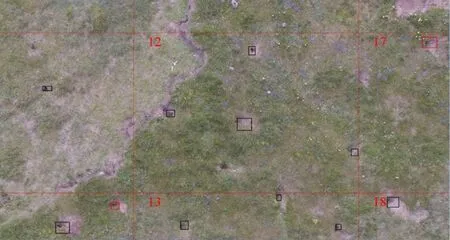
图4 航拍照片分析鼠兔洞口数量示意图Fig. 4 Diagram showing analysis of the number of pika burrows with aerial photographs
采用单因素方差分析法分析不同样地类型鼠兔洞口密度和洞道特征,并采用线性回归分析雷达获取的波动数量与高原鼠兔洞道的相关性。所有数据采用Excel 2013版、SPSS 20版、matlab R2017a统计软件进行图表制作和统计分析。
2 结果
2.1 不同样地类型高原鼠兔的洞道结构
不同样地类型高原鼠兔洞道直径约为6 cm。高原鼠兔洞道内部结构复杂程度不一,不同植被条件下洞道结构差异明显,但整体呈现近洞口和高原鼠兔洞道末端处较浅,洞道中部深度较深的“凹”形结构,平均最深深度可达地下40 cm左右;当洞道末端没有出口时,洞道末端距离地面约为16 cm。就复杂程度而言,退化沼泽高原鼠兔洞道最为复杂(图5),其进出口有2个,在洞道最深处挖出1个环形通道;就长度而言,样本中最短的鼠兔洞道仅有85 cm,图中左边洞口,末端在地面以下16 cm处;最长的鼠兔洞道长度超过560 cm,左端为一个洞口,进入之后有深度的起伏,在右端一个横断面又出现一个洞口。
2.2 不同样地类型高原鼠兔洞口密度及洞道特征
高寒草原洞口数和洞口密度均高于其他样地类型,退化高寒草甸洞口数和洞口密度最低(表1)。不同样地类型探地雷达探测高原鼠兔洞道产生的波动数不同(表1),其中退化高寒草甸平均波动数最高为3.1个,高寒草甸次之,为2.8个,退化沼泽和高寒草原分别为1.9和1.7个。根据探地雷达的工作原理和平均波动数的来源,高寒草甸和退化高寒草甸高原鼠兔洞道的平均长度显著长于高寒草原与退化沼泽(P<0.05),这与实地测定的结果相符。
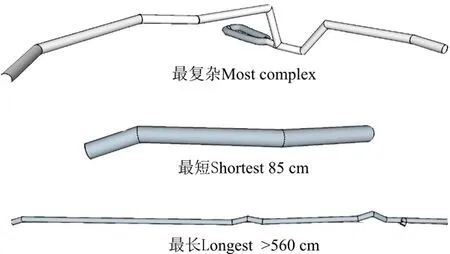
图5 3种典型高原鼠兔洞道结构三维图Fig. 5 Three-dimensional pictures of three kinds of typical tunnel structures of plateau pika
2.3 实地探测数据与雷达数据的相关性
通过对同一个样方内的探地雷达测量的高原鼠兔洞道的波动数与实际洞口数回归分析发现,二者具有显著的线性相关(R2=0.54,P<0.01)(图6);探地雷达测量的波动数据/实际的洞口数量与平均洞道长度的线性拟合(图6)结果表明,二者之间也呈极显著相关性(R2=0.64,P<0.01)。

表1 不同样地类型高原鼠兔洞道结构特征Table 1 The structural characteristics of plateau pika burrows
同列不同小写字母表示差异显著(P<0.05)。
Different lowercase letters within the same column indicate significant difference at the 0.05 level.

图6 波动数量与高原鼠兔洞道关系Fig. 6 Relationship between the fluctuations and the number of plateau pika tunnels
3 讨论
受观测手段和研究方法的限制,对于高原鼠兔洞道结构的研究通常采用实地挖掘的方法[18],具体步骤为从洞口处沿着洞道方向将洞道上方的土壤和植被移开,以便测定洞道直径、深度、拐点、窝巢等特征。也有学者采用化学材料固定洞穴结构对小型哺乳动物的洞穴进行研究[19]。这些方法不仅耗时费力且对生态系统环境具有破坏性,而且会引起洞穴的坍塌,还存在无法获取足够的数据量的问题[7]。本研究通过实地调查,对不同植被类型高原鼠兔洞道进行剖析,首次采用探地雷达对高原鼠兔洞道结构进行研究,并通过实际观测的结果进行了验证。从实地调查的结果来看,不同植被类型的高原鼠兔洞道结构有一定差异,高寒草甸植被和退化高寒草甸高原鼠兔洞道的平均长度显著长于高寒草原与退化沼泽(P<0.05)。这可能与鼠兔栖息地环境以及食物来源有关,郭新磊等[20]的研究表明,植被和土壤水热是影响高原鼠兔选择栖息地的主要因子,高原鼠兔偏向于选择植被条件较好、土壤含水量较高,生长季陆地表面温度较低的区域。退化沼泽鼠兔洞道结构最为复杂,与前人的研究结果[7]一致。沼泽草甸可以为高原鼠兔提供充足的食物来源,但是沼泽草甸土壤水分较高,尤其降水事件发生后存在洞道被淹的风险。因此,高原鼠兔选择水分相对较少的退化沼泽作为栖息地,但是仍然可能面临水淹的风险,可能为避免洞道被冲毁,经过长时间的挖掘产生了大量分支[21]。
探地雷达测量的高原鼠兔洞道波动数与实际洞口数、平均波动数与平均洞道长度均具有较好的相关性(图6),这与冯温雅等[11]应用便携式探地雷达,用主成分分析(PCA)法在草地进行试验得到的结果一致,其研究表明探地雷达在高原鼠兔洞道的探测具有可行性。但是在实际测量中,由于雷达波动数据是在10 m×10 m的样方内测量的所有高原鼠兔洞道波动的数据,而高原鼠兔洞道的长度数据是多个高原鼠兔洞道的平均值,二者不能直接用来比较分析。本研究将样方内平均每条鼠兔洞道的波动数与洞道长度进行拟合,二者呈现较好的线性关系(图6),说明探地雷达在高原鼠兔洞道探测的结果与实际观测的结果相符,在高原鼠兔洞道的探测方面具有一定的适用性和可行性。该方法工作量小,无损快速,可用于大范围高寒草地高原鼠兔洞道结构的调查研究。但在实际应用过程中,探地雷达可能会对同一个洞道进行重复测量,导致出现多个波动。此外,由于地面的凹凸不平,即使使用测距轮测量的方式,在测量过程中也难免出现天线系统上下颠簸,致使雷达的探测结果出现小部分的图像不连续,增加了后期处理的难度。本研究区土壤富含砂砾石,部分砂砾石大小和鼠兔洞道直径相近,在雷达探测时也会显示波动,增加了雷达波动的数量。因此,在将来的研究工作中还要进一步探索,寻求改进的方法。
4 结论
本研究采用实地观测的方法调查了典型高寒草地高原鼠兔的洞道结构,并以实测数据验证了雷达在探测鼠兔洞道结构的可行性与适用性。研究结果发现高原鼠兔洞道的直径约为6 cm;高原鼠兔洞道内部结构复杂程度不一,不同植被条件下洞道结构有显著差异,但整体呈现近出口(或无出口洞道末端)和入口处较浅,洞道中部深度较深的“凹”形结构,平均最深深度可达地下40 cm左右;当洞道末端没有出口时,洞道末端距离地面厚度约为16 cm。就复杂程度而言,退化沼泽高原鼠兔洞道最为复杂;长度从85~560 cm不等;探地雷达测量的高原鼠兔洞道波动数量与实际测量高原鼠兔洞口数量以及平均波动数量与高原鼠兔洞道长度均具有较好的相关性。本研究结果表明,探地雷达在高原鼠兔洞道的探测方面具有一定的适用性和可行性,且该方法无损快速,为大范围调查高寒草地鼠兔洞道结构提供了一种新的方法。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
房地产导刊(2022年10期)2022-10-18
青海草业(2022年2期)2022-07-23
建材发展导向(2021年16期)2021-10-12
科技研究·理论版(2021年20期)2021-04-20
雷达学报(2021年1期)2021-03-04
活力(2019年21期)2019-04-01
通信电源技术(2018年3期)2018-06-26
河北林业科技(2016年5期)2016-11-08
中国工程咨询(2016年1期)2016-02-14
