
温性草原利用方式对生态系统碳交换及其组分的影响
2018-12-20 06:43李愈哲樊江文胡中民邵全琴
生态学报 2018年22期
李愈哲, 樊江文, 胡中民, 邵全琴
1 中国科学院地理科学与资源研究所陆地表层格局与模拟重点实验室,北京 100101 2 华南师范大学地理科学学院,广州 510631
人类活动造成的大规模土地利用、覆盖变化(LUCC)及其生态影响是全球变化领域的重要研究热点[1-2]。当前,内蒙古中东部温性草原区域受到气候变化和人类活动的影响,土地管理利用格局变化剧烈[3]。其特殊的气候环境和生态条件又使得区域内生态系统对土地管理利用变化的响应尤为敏感、迅速[4],成为研究LUCC生态效应的理想区域之一。
天然草地放牧是内蒙古中东部温性草原区域最为主要的土地利用方式[5],农田原本较多分布在东南部灌溉条件较好或局部地形小气候更为湿润的区域。受气候变化驱动,农牧交错带逐渐北移[6],草原区域的局地适耕性水平快速提高[7]。随着国内经济、人口高速增长,人类对粮食资源和生活空间的需求快速增大。自1949年以来我国共开垦草地1930万hm2,占我国现有草地面积的4.8%,全国现有耕地的18.2%源于草地开垦[8]。截至2000年,仅内蒙草原东部区域的开垦面积就超过1000 km2[9]。草原区土地开垦利用呈逐渐蔓延扩展之势。另一方面,自20世纪80年代至2002年,我国牧区草地载畜压力不断增加,草原区域承载牲畜头数增加了46%[10]。随着放牧利用强度的非理性增长,草原放牧区域面临着载畜压力增大、草场过牧退化等生态问题[11- 13]。为应对日益严峻的草地生态环境问题,各级政府不断加大投入,在草原区实施了系列生态恢复工程。随着工程的深入建设实施,以围封禁牧为代表的草原恢复方式应用面积逐年扩大,其中截止2006年底仅京津风沙源治理工程实施的围栏封育面积就达到66.93×104hm2[14]。草原区域内大面积的开垦利用,以及草地的长期禁牧恢复将对原本放牧利用的生态系统的产生哪些影响亟待深入的了解与研究[15-16]。
已有研究证实土地管理利用方式变化在欧洲山地和美洲普列利草地能对草地生态系统碳交换(Net ecosystem exchange, NEE)造成巨大影响[17-18]。NEE是植被总初级生产力(Gross primary productivity, GPP)与总生态系统呼吸(Ecosystem respiration,Re)之和,而Re仍存在复杂的组分来源[19-20]。了解NEE中组分的具体变化如何最终贡献NEE的改变对于机理性了解生态系统NEE的调控非常重要[21]。生态系统呼吸依据释放来源的空间差别,可以区分为地下部分土壤呼吸(Below ground respiration,Rb)和地上部分植物体呼吸(Above ground respiration,Rab)两部分。而生态系统呼吸根据CO2释放来源生物属性,又常被拆分为来自植物体的生态系统自养呼吸(Autotrophic respiration,Ra)和主要释放自微生物和土壤动物的异养呼吸(Heterotrophic respiration,Rh)。异养呼吸和自养呼吸的准确拆分往往被认为是植被NPP测算的关键[22]。土壤呼吸依据释放来源的不同,又可细分为3个生物学过程(土壤微生物呼吸、根呼吸和土壤动物呼吸)和一个非生物过程(含碳物质化学氧化作用)[23-24]。其中土壤呼吸中含碳矿物质的化学释放一般情形下非常弱[25],对土壤呼吸的贡献极小,此外以往研究表明土壤动物呼吸所贡献的比例也不高[26-28]。因此,土壤呼吸的区分目前主要关注根系呼吸(Root respiration,Rr)和土壤微生物呼吸的准确拆解[29]。呼吸过程是地表向大气输送温室气体的最主要方式,深入细致的了解呼吸及其组分对人类典型土地管理利用方式的响应规律是了解区域和全球碳平衡的一个重要环节[30]。
开垦利用和禁牧恢复作为我国温性草原区域最具代表性的土地管理利用方式转变,研究其对放牧草地生态系统物质交换的影响,将有助于预测草地管理利用方式的大规模转变如何影响生态系统碳收支和区域物质平衡,从而帮助准确评估土地管理利用转变的生态效应和气候影响。本研究利用人工处理控制实验和野外观测相结合的方式,针对温性草原区域3种典型的土地管理利用方式放牧、禁牧和开垦农用,测算了生态系统碳交换及其精细组成,旨在准确了解土地管理利用方式变化如何影响区域地表与大气间碳交换,同时尝试机理性解析光合、呼吸过程中哪些组分在管理利用方式间的变异贡献了碳交换的改变,探讨其变异原因。
1 材料与方法
1.1 样地介绍
实验选取锡林郭勒大针茅+糙隐子草草地(Form.Stipagrandis+Cleistogenessquarrosa)植被类型。据气象资料(1982—1998),区域年平均气温0.6℃,最冷的1月和最暖的7月月平均气温分别为-21.4℃和18.5℃。年均降雨量350 mm,主要集中在6—8月,年内多大风,气候干燥。土壤类型以栗钙土为主[22]。天然草地自由放牧是当地最主要的土地利用方式。局地水热条件亦可适宜农作物生产[31-32],草地开垦农用现象突出。本实验选取3块毗邻且利用/管理方式不同的草地。除选择典型放牧草地(Grazing, G)外,西北0.8 km为1979年开始的围封禁牧样地(Grazing excluded, GE),地表有较多凋落物。在G东侧1.5 km处为开垦的农用样地(Reclaimed, R),原生植被类型与G相似,2010年开垦翻耕种植农作物。在典型放牧草地中心部(43°32′24.06″N,116°33′41.76″E)设有涡度相关系统,用以监测植被-大气间水碳热交换通量,通量塔与GE、R样地边界均超过距离800 m以上。
1.2 实验方案
实验观测采用巢式随机区组设计,依照巢式设计在毗邻的G、GE和R的3个样地内,分别设立4个试验区组作为重复(区组间间隔超过5 m)。每个区组内部包含4个1 m×1 m样方,样方之间间距1.5 m,以避免彼此影响。每个区组随机选取1个样方用于地上、地下生物量测定及LAI、土壤取样,方法见[33]。另3个样方随机分配以3种不同的处理方式,以测定和拆分生态系统碳通量及其组分。在测定开始两周之前,每个区组内随机选取一个样方喷施除草剂(41%草甘膦异丙铵盐水剂稀释150倍),2 d后该样方内植物全部被杀死并呈枯立起状保留在样方内,维持样方植物存活时自然的遮阴情形(Dry and stand,DS)。区组内另一随机样方,与以往测定生态系统蒸发速率的处理一致,齐地面刈割地表植被[22,34],并在每次测定前一天剪除其地面新萌出部分,维持样方地表裸露状态(Clipping, CL)。剩余的一个样方不做处理维持原状,设为空白对照(Control with plants,C)。在G、GE和R的3个草地利用方式样地内共计设置48个样方,每个样地中对照样方、刈割处理样方、除草剂处理样方均重复4次(分置于各样地的4个实验区组中),剩余每个样地的4个区组重复中各留有1个收割样方用于相关群落特征测定。在空白对照样方内利用箱式法测得的NEE结果与采用标准校正处理[35]后的涡度相关系统测得NEE进行了比对,两者具有很好的一致性(图1)。
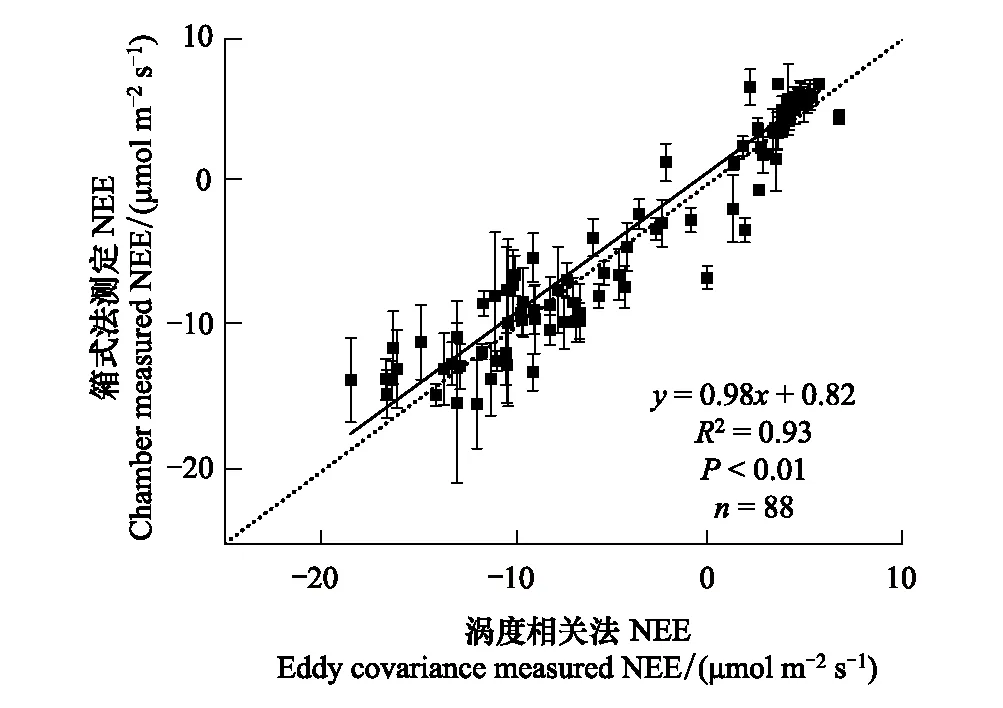
图1 放牧样地NEE同化箱测定法和涡度相关测定法结果间对比Fig.1 Comparison of the ecosystem carbon exchange obtained by the chamber method and eddy covariance systemNEE: 净生态系统交换Net ecosystem exchange
1.3 生态系统碳交换测定及拆分计算
对照样方用于测定生态系统CO2净交换速率(NEE),同时借助黑箱遮光法,测定生态系统呼吸速率(Re)。DS用于测定生态系统异养呼吸速率(Rh),刈割样方CL内测定的CO2可视作生态系统地下部分的呼吸速率(Rb)。
每个测定样方中,在测定的前两周安置一个正方形金属底座(面积50 cm×50 cm,高10 cm)。底座下边缘锋利,方便压入土中,同时留有3 cm高度高出土面以便紧密连接同化箱。生态系统水碳交换中前述组分(NEE,Re,Rh和Rb)都借由一个立方体同化箱(50 cm×50 cm×50 cm)连接一台红外气体分析仪(LI- 6400, LI-COR Inc., Lincoln,NE, USA)在相应处理样方直接测定,记录测定方案如Niu等描述[35]。两枚小型风扇安装在同化箱内顶部,用以在测期间混合空气,一个温度探针被放置在同化箱中心测定空气温度,一个土壤温度探针被插入同化箱附近土壤中用于测定5 cm深处土壤温度,同时利用TDR100测定了土壤体积含水。进行测定时,透明的同化箱紧密放置在金属底座上,为防止密封不严的情况,在连接处安装了橡胶密封条。同化箱被合适的放置在底座上后,LI- 6400根据输入程序,自动连续测定箱内CO2浓度30 s,每秒记录一次。数据从头开始每10个点进行线性拟合,当R2>0.98时即接受斜率用以推算CO2通量,否则移动至2—11点重复,依次类推直到取到符合要求的斜率。
观测在草地的最大生物量时期(8月)选择晴朗、微风,天气条件相近时在不同利用方式草地交叉密集进行,共测定12个全天(每利用方式4个重复观测日)。对各样方生态系统碳交换的观测每隔1 h测定一轮,连续进行24 h。G样地测定结果经过与涡度相关系统测水碳通量的比较,两个测定方法的结果具有良好的一致性,测定是准确可靠的(图 1)。
生态系统碳通量直接测定计算的方法在以往研究中有详细描述[22,35],本研究中对通量各组分日积累量的计算公式如下:
对于生态系统呼吸及其各组分在内的碳释放过程,因为交换速率全部为正值,可直接通过交换速率进行计算,
式中,Fc是碳通量(μmol m-2s-1; 负值Fc代表生态系统吸收固定大气中的CO2,反之是生态系统释放CO2到大气)。
对于生态系统的碳吸收过程(GEP、NPP、NEP),因各组分交换速率多为负值,为方便对其日积累量进行直观比较,计算方式为,
生态系统CO2通量收支中的其他一些组分如总生态系统生产力(GEP),自养呼吸速率(Ra)等则可通过差值法[36],计算如下。
GEP=NEP +Re
净生态系统生产力NEP是净初级生产力(NPP)中去除土壤动物和土壤微生物的异养呼吸Rh后的剩余部分,由此:
NPP=NEP +Rh
生态系统呼吸Re由植被本身自养呼吸消耗Ra和生态系统异养呼吸Rh共同构成,利用测得的Re和Rh,自养呼吸Ra依据下式求得:
Ra=Re-Rh
从空间角度考虑,生态系统呼吸由地上呼吸(Rab)和地下呼吸Rb两部分组成。利用测得的地下部分呼吸Rb和生态系统呼吸Re,生态系统地上部分呼吸Rab计算如下:
Rab=Re-Rb
其中地下部分呼吸Rb,在根系正常活动时,即可视作土壤呼吸(Rs),包含土壤微生物呼吸、根系呼吸和土壤动物呼吸3个生物学过程,以及含碳矿物质的化学氧化等非生物CO2释放。在大部分土壤类型中和绝大多数常见生态系统中,土壤含碳矿物质化学氧化和动物呼吸所释放的CO2所占比例极小,可以忽略不计。由此,土壤呼吸主要由两大来源组成,即土壤中的微生物呼吸和土壤中根系呼吸。土壤微生物异养呼吸占据生态系统异养呼吸Rh的绝大部分。生态系统自养呼吸Ra,依据空间差异,由生态系统地上部分植被呼吸Rab和地下根系Rr共同构成,由此可推导隶属于Ra的一部分的根系呼吸可计算如下:
Rr=Ra-Rab=(Re-Rh)-(Re-Rb)=Rb-Rh
生态系统碳交换各组分的日交换量通过对24小时通量速率进行全天积分获得。
1.4 生态系统碳释放组成
生态系统呼吸空间上由地上部分呼吸Rab和地下部分呼吸Rb通量之和,生态系统土壤呼吸占生态系统呼吸的比例fsoil:
fsoil=Rb/Re
生态系统呼吸来源上,由植物体产生的自养呼吸Ra和异养生物产生的异养呼吸Rh共同构成,生态系统自养呼吸占生态系统呼吸fa:
fa=Ra/Re
土壤呼吸通量等于根呼吸Rr和土壤微生物呼吸Rh之和,根呼吸占土壤呼吸的比例froot:
froot=Rr/Rs=Rr/Rb
根呼吸站生态系统呼吸的比例froot_re:
froot_re=Rr/Re
1.5 数据分析
使用配对t-检验分析碳交换及其组分日动态差异,日交换量采用HSD法单因素方差分析(One-way ANOVA)进行检验。呼吸组成与环境因子间关系分析采用线性回归法。所有数据分析使用了SPSS 12.0,作图采用Originpro 8.0。
2 结果与分析
2.1 不同利用方式草地光合呼吸各组分日动态和交换量
3种管理利用类型的草地,NEE、GEP和NPP速率的日动态大致都呈倒U型曲线(图2)。白天3种利用方式生态系统的NEE彼此间差异不大,但夜间开垦区域的碳排放要明显高于放牧和禁牧草地(图2)。GEP在3种不同利用方式草地在12:00—13:00间均出现不同程度的午休现象(图2)。配对T检验结果显示,3种不同利用方式草地白天NPP水平彼此间差异不大(P>0.05)。
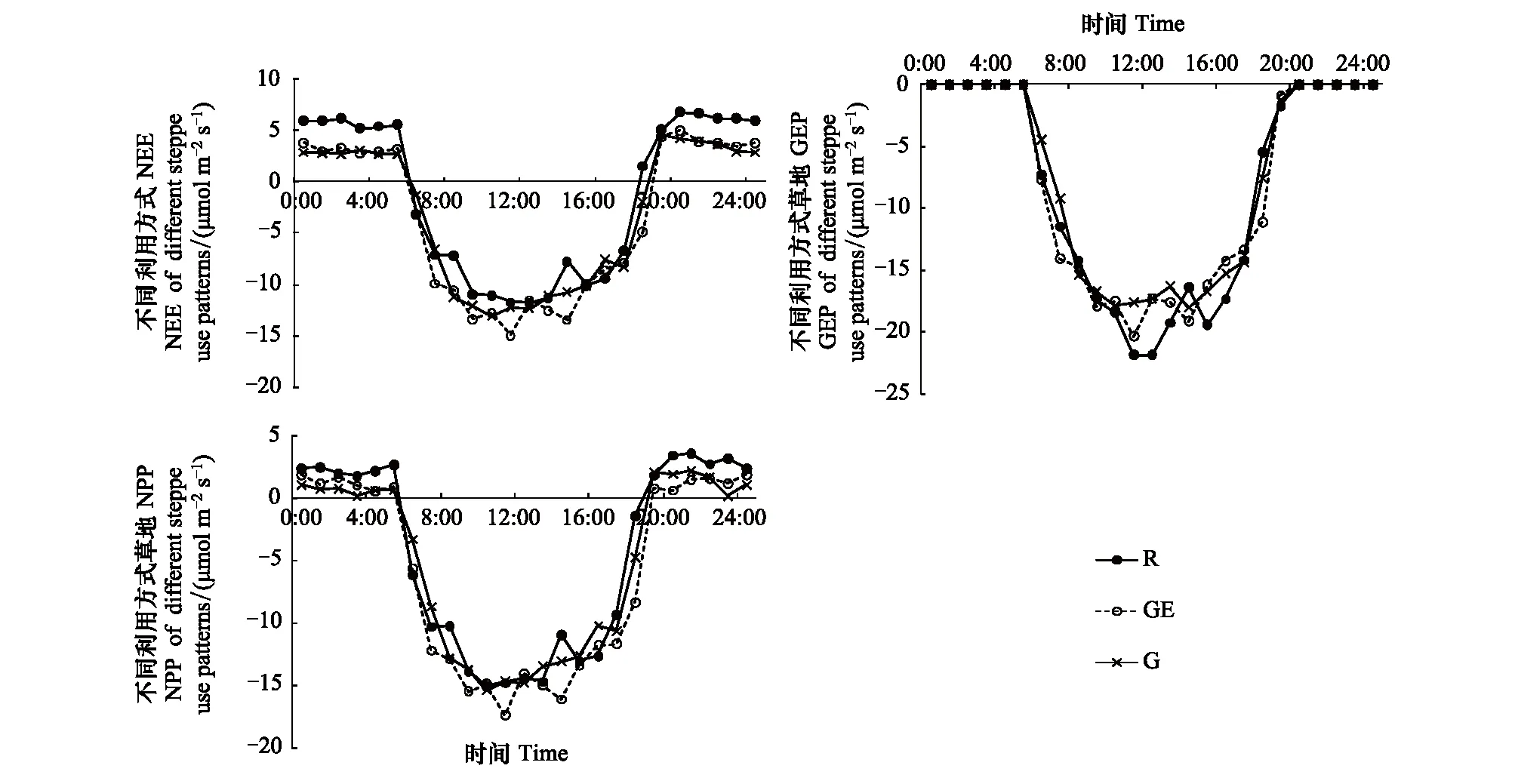
图2 不同利用方式草地光合过程日动态Fig.2 Changes of daily dynamics of NEE, GEP and NPP in different grassland use pattern during the peak growing period GEP: 总生态系统生产力 Gross ecosystem productivity; NPP: 净初级生产力Net ecosystem primary productivity; G:放牧 Grazing; GE:禁牧 Grazing excluded; R:开垦 Reclaimed
生态系统呼吸在不同利用方式草地均呈峰值出现在午后13:00—16:00的单峰曲线,放牧和禁牧样地两者间差异不大,在各时段释放CO2均低于开垦样地的呼吸水平(图3)。开垦样地的自养呼吸在各时间节点均高于放牧和禁牧草地,峰值出现在12:00, 放牧和禁牧草地Ra峰值出现14:00(图3)。异养呼吸的日动态则相对平稳,Rh在3种利用方式草地间的差异也较Ra小(图3)。开垦样地的地上和地下部分呼吸均强于放牧和禁牧草地(图3)。在根系呼吸方面,不同利用方式样地根系呼吸最大值均出现在开垦样地午后13:00,接近2 μmol m-2s-1,此外在日出后的7:00—10:00间,开垦样地的根呼吸出现负值,根系呈现出吸收CO2的迹象,这一现象在放牧和禁牧草地并没有出现。
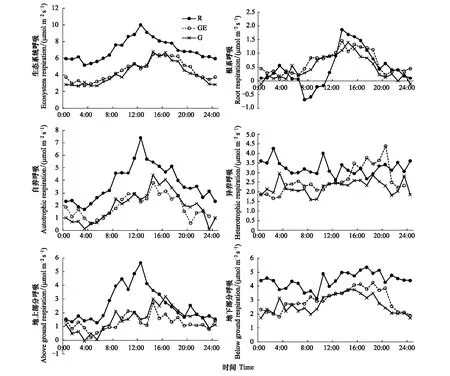
图3 不同利用方式草地呼吸各组分日动态Fig.3 Changes of daily dynamics of ecosystem respiration compositions in different grassland use pattern during the peak growing period
3种利用方式草地的GEP水平大致位于30—35 g CO2m-2d-1。与放牧草地GEP相比,开垦和禁牧均趋向于增加生态系统GEP(图4),其中开垦样地显著增加12.7%(P<0.05)。然而草地开垦显著降低原有放牧草地的NPP(13.0%,P<0.05),而长期禁牧草地与放牧草地NPP彼此间差异不大(P>0.05)。草地里管理利用方式差异对NEP影响情形与NPP相似,R显著低于G和GE (P<0.05)。但开垦对放牧草地NEP的降低幅度较NPP大,达到51.2%。
放牧和禁牧草地Re彼此间差异不大(P>0.05),均在17 g CO2m-2d-1左右(图5)。草地开垦后的Re达到26.9 g CO2m-2d-1,较放牧草地显著增加64.3%(P<0.05)。放牧和禁牧草地Ra彼此间差异不大(P>0.05),均在8 g CO2m-2d-1左右。草地开垦后的Ra达到14.1 g CO2m-2d-1,较放牧草地显著增加88.8%(P<0.05)。放牧和禁牧草地Rh彼此间差异不大(P>0.05),均在9 g CO2m-2d-1左右。草地开垦后的Rh达到12.8 g CO2m-2d-1,较放牧草地显著增加43.7%(P<0.05)。放牧和禁牧草地Rab彼此间差异不显著(P>0.05),而草地开垦后的Rab显著增加为放牧草地的两倍左右(P<0.05)。放牧和禁牧草地Rb彼此间差异不大(P>0.05),均在11 g CO2m-2d-1左右。草地开垦后的土壤呼吸升至14.6 g CO2m-2d-1,较放牧草地显著增加了35.6%(P<0.05)。根系呼吸水平在3个不同利用方式间差异不显著(P>0.05),均在2 g CO2m-2d-1左右。
2.2 不同利用方式草地呼吸组成及其控制因素
在3种不同管理利用方式下,土壤呼吸占生态系统呼吸比例都超过50%,禁牧草地土壤呼吸贡献比例最大,达到67%。与放牧草地相比,开垦农用显著降低土壤呼吸占生态系统呼吸比例,下降幅度为17.7%(P<0.05)。而围封禁牧与放牧草地之间无显著差异(P>0.05)。生态系统呼吸中自养呼吸Ra所占比例在不同草地管理利用方式下彼此间差异较小,都在50%左右。其中开垦农用Ra贡献比例最大(52.3%),相比最低的放牧草地(44.5%)增幅为17.5%(P<0.05)。根系呼吸对土壤呼吸的贡献比例在3个样地之间差异相对较大。与放牧草地相比,草地开垦农用显著降低了froot(表1,P<0.05)。而围封禁牧则趋向于增加草地中根系呼吸对土壤呼吸的贡献比例,与放牧草地相比增幅为3.7%,但两者间差异未达显著水平(P>0.05)。与放牧草地相比,开垦农用显著降低根系呼吸占生态系统总呼吸的比例(表1,P<0.05),然而froot_re在围封禁牧与放牧草地之间无显著差异(P>0.05)。

表1 不同管理利用方式下3种草地生态系统呼吸、土壤呼吸拆分结果
均值± 标准误;同行内不同小写字母代表相互间差异显著(P<0.05);Re:生态系统总呼吸, ecosystem respiration;Rs: 土壤呼吸, soil respiration;Ra:自养呼吸, autotrophic respiration;Rr,根系呼吸, root respiration;fsoil: 土壤呼吸占Re比例, the ratio ofRstoRe;fa:自养呼吸占Re比例, the ratio ofRatoRe;froot: 根系呼吸占Rs比例, the ratio ofRrtoRs;froot_re: 根系呼吸占Re比例, The ratio ofRrtoRe
在众多环境因子中,各样点间地下生物量变化表现出与fsoil极显著正相关关系,其改变能95%程度上解释fsoil在样点间的变化(图6,P<0.001)。而样点间LAI差异则解释了样点间fa变化的65%,LAI在样点间增加与fa变化呈正比(P<0.001)。土壤呼吸中根系呼吸贡献比例froot表现出与地下生物量的较好相关性(P<0.001),地下生物量增加可以解释froot增加的77%。生态系统总呼吸中根系呼吸贡献比例froot_re与总生物量间呈显著正相关关系(P<0.001),总生物量增加可以解释froot_re变化的84%(图6)。

图4 不同利用方式草地日均GEP、NPP和NEPFig.4 Mean daily GEP, NPP and NEP in different grassland use/manage pattern during the peak growing period 均值±标准差; 同组中不同小写字母代表相互间差异显著(P<0.05)

图5 不同利用方式草地呼吸各组分日交换量 Fig.5 Mean daily ecosystem respiration and its components in different grassland use/manage pattern during the peak growing period 均值±标准差; 同组中不同小写字母代表相互间差异显著(P<0.05)
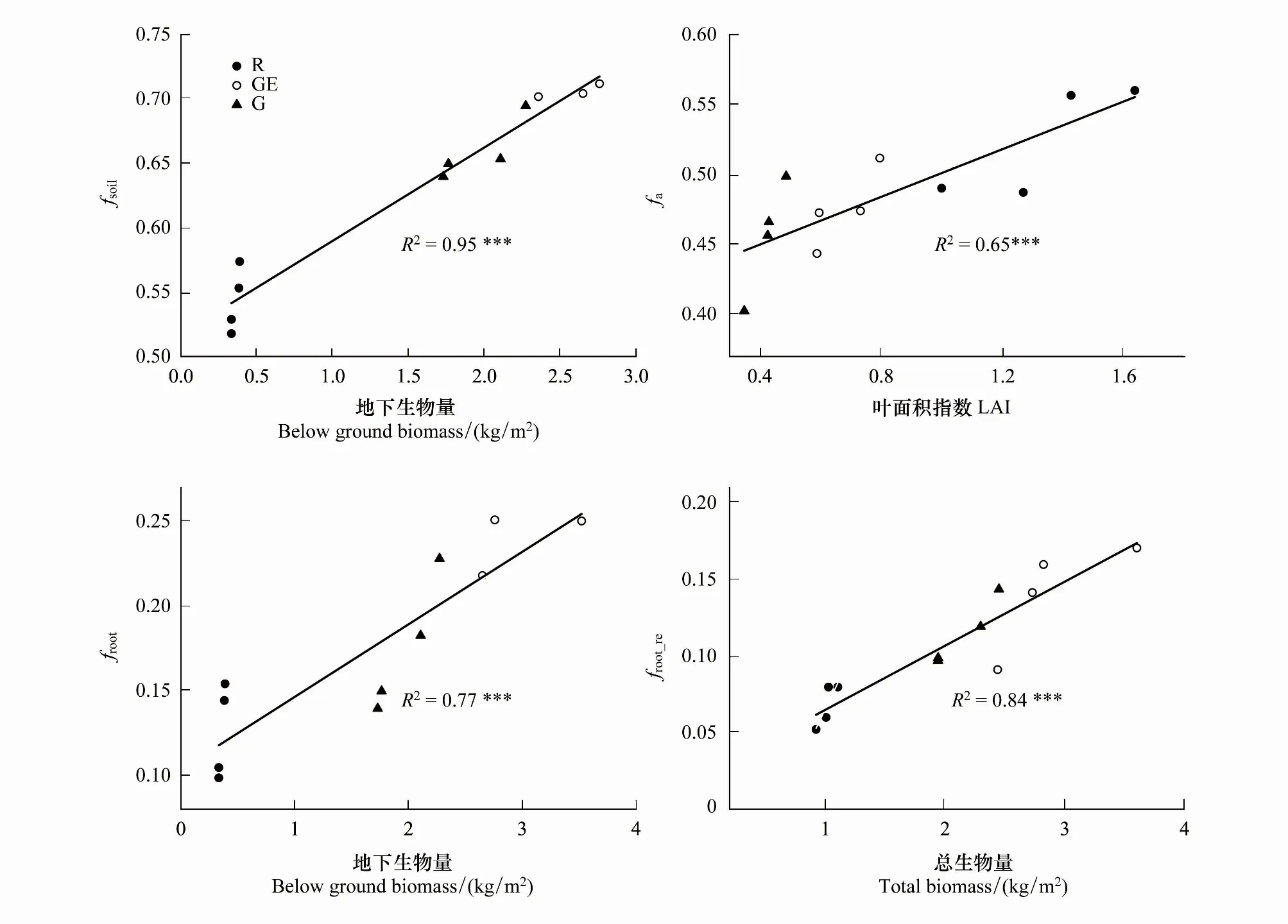
图6 不同管理利用方式草地fsoil, fa, froot和与froot_re群落环境因子间关系Fig.6 Relationships between fsoil, fa, froot and froot_re on below ground biomass, leaf area index (LAI),total biomass across the three different grassland use/management pattern fsoil: 土壤呼吸占Re比例 The ratio of Rs to Re; fa:自养呼吸占Re比例 The ratio of Ra to Re; froot: 根系呼吸占Rs比例The ratio of Rr to Rs; froot_re: 根系呼吸占Re比例The ratio of Rr to Re.*, P<0.05; **, P<0.01; ***, P<0.001
2.3 不同利用方式草地生态系统根系呼吸活力与根冠比关系
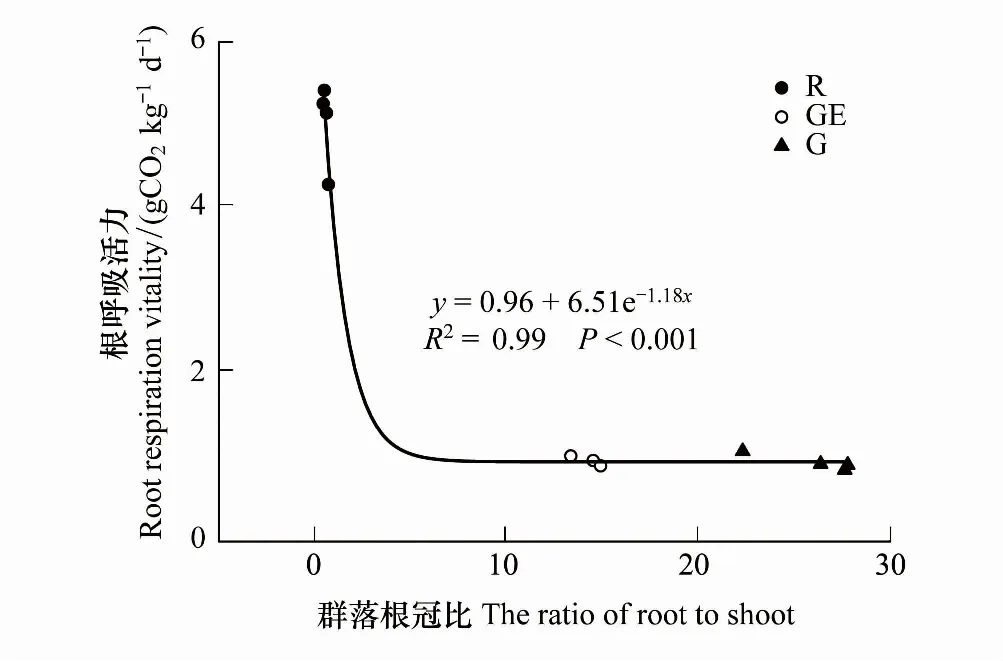
图7 根呼吸活力和根冠比的相关关系 Fig.7 Correlation between root respiration vitality and the ratio of root to shoot biomass
根呼吸活力被定义为单位根生物量的呼吸速率[37],根据Rr与活根生物量之比,可计算出根呼吸活力。3种不同草地管理利用方式下,植被根呼吸活力变化范围在0.86—5.4 g CO2kg-1d-1之间。根呼吸活力在放牧和禁牧草地之间差异并不显著(P>0.05),而开垦为农田后根呼吸活力大幅上升(图7,P<0.05)。在草原区域不同管理利用方式下,与史晶晶等[38]研究结果相似,植被根冠比与根呼吸活力间表现出极显著的负指数相关关系(图7,P<0.001),解释程度为99%。
3 讨论
3.1 土地管理利用方式改变对草地碳交换的影响
本研究中放牧草地和禁牧恢复草地,都具有比开垦区域显著更高的NEP。表明大规模的开垦行为,不仅改变生态系统的物种组成和群落结构[22, 39],还可能弱化北方草原区域天然草地生态系统的原有的固碳能力。尽管毗邻区域不同利用方式生态系统间,GEP彼此间差异相对较小(图2、图4)。这体现了植被将光合有效辐射转化成GEP的能力往往差异不大,这一特性应用在了若干光能利用效率为基础的生产力模型中[39]。开垦区域的NPP相比天然草地(包括放牧草地和禁牧草地)下降约13%(图4),而开垦区域NEP相比两天然草地下降了约51%(图4),NEP的降幅远高于NPP。这反映出单纯模拟NPP变化可能不能充分、准确评估区域生态系统碳固定能力在土地管理利用变化影响下的损失。只考虑NPP变化将会低估草地开垦可能造成的区域碳固定下降。
具有较高GEP的开垦样地,NPP和NEP反而显著低于放牧和禁牧草地(图4)。这一情形应归咎于生态系统呼吸及其组成在利用方式转变下发生了巨大的改变[40]。如开垦样地显著较低的NEP就归咎于增加幅度超过50%的生态系统总呼吸(图5,P<0.05)。开垦区域Re的显著上升可能主要归因于小麦作物自身的高代谢[41]使得生态系统自养呼吸强度大幅度高于同区域的天然草地(P<0.05),这一变化也造成开垦样地NPP显著低于放牧和禁牧草地。另一方面,由于农田施用农家肥使得土壤中微生物呼吸产生Rh大幅增强(P<0.05)。最后,虽然草地开垦极大的减少了群落的地下生物量(即根系数量)[33],但是由于根冠比下降导致的根系呼吸活力指数增加[38](图7),农田整体根系呼吸并没有出现显著下降(图5,P>0.05)。草地开垦区域生态系统增加的Rh和Re如果长期持续,无疑将逐渐削弱区域的土壤碳库,增加区域的CO2释放。而草原区域大规模的开垦,通过碳收支的改变,无疑将对区域乃至全球气候产生一定程度的暖化贡献。
3.2 土地管理利用方式改变对草地呼吸组成的影响
通过对比不同管理利用方式草地碳交换及其组成,生态系统呼吸及其组分的变化应是影响温性草原区域生态系统最终NEP的重要因素。本研究中,自然物种组成的草地生态系统(包括放牧和禁牧草地),其总呼吸中土壤呼吸占比fsoil彼此接近,均显著高于草地开垦后的生态系统(表1)。这主要由于开垦后植被地上部分生物量比例大幅增加造成地上部分呼吸贡献的极大增加,反映出草地开垦后地上地下生物量分配的巨大改变[33]。相比fsoil,自养呼吸在生态系统呼吸中的占比fa在不同管理利用方式间的差异较小且更具连续性,禁牧草地与具有较高fa放牧草地以及与较低fa开垦草地间差异都不显著,3种群落的fa大致都在50%左右。不同草地利用方式下,较高的地上部分生物量可预期的将趋向贡献较多的自养呼吸[42],从而使生态系统表现出较高的fa。史晶晶等在邻近羊草草地[38]及李凌浩等[43]在邻近区域放牧草地测得的froot都在24%左右,略高于我们的测定结果,这一结果与本研究围封禁牧草地测得的froot21.2%非常接近,表明放牧活动和草地开垦趋向于降低群落中的根系呼吸所占比。考虑到羊草草地的根系生物量[38]与围封禁牧草地非常接近[33],远高于放牧和开垦区域。因此相比禁牧草地和羊草草地[38],开垦样地和放牧样地较低根系呼吸贡献率推测是由于地下生物量及其占比更低所致。
生态系统地下生物量的增加能很好解释土壤呼吸对生态系统呼吸贡献的上升(图6,95%)。地下生物量与fsoil表现出的极显著正相关可归因于根际激发效应[25]。以往研究还指出地下生物量的增加不仅趋向于直接增加根系的呼吸[44],增加的根系还通过释放根际分泌物和改变土壤理化性质可以促进土壤有机质的分解,从而加强微生物呼吸。自养呼吸贡献比例与LAI表现出极显著正相关(图6),反映出了LAI与产生自养呼吸的植物生物量所隐含的正相关关系。更高的LAI往往对应更高的植物活体生物量[45-46],从而表征更高Ra及其比例。froot与根系生物量呈现出极显著正相关关系(图6),根系的增加促进根系呼吸在Re中所占比重。然而围封禁牧草地根系生物量是毗连开垦放牧草地的10倍,而froot只增加了10%左右。这固然关系到不同群落建群种本身固有特性,但也可部分归因于前述的根系激发效应造成土壤微生物呼吸增加[25]。此外,如图7所示,根系呼吸效率与根冠比间呈极显著的负指数关系,开垦区域较少的土壤根系生物量往往具有极高的根系呼吸效率,从而仍然能产生出不低的根系呼吸,近似现象在以往研究中曾被提及[38]。
4 结论
研究结果表明,温性草原区域的草地管理利用方式转变能显著的改变生态系统的碳交换水平,并大幅改变碳交换的组分构成。其中草地开垦成农田对生态系统碳交换的影响最为强烈,主要表现为NEP的显著下降(51.2%)。造成这一现象的原因是种植作物的自养呼吸水平Ra相较天然植被大幅提高(增加近90%),同时施肥后的土地土壤异养Rh也显著增加(43.7%),最终造成生态系统总呼吸Re相比放牧草地显著增加64.3%。长期禁牧也趋向减少生态系统NEP,但与放牧草地相比,变化不显著(10.0%,P>0.05)。相比长期禁牧,草地开垦将严重弱化区域植被的碳固定作用,长远看可能对草原区域植被碳库和土壤碳库的积累产生负面影响。草地开垦区域生态系统Rh和Re的增加如果长期持续,将逐渐削弱区域的土壤碳库,增加区域的CO2释放。而草原区域大规模的开垦,通过碳收支的改变,将可能对区域乃至全球气候产生一定的暖化作用。
猜你喜欢
林业资源管理(2022年6期)2023-01-18
林业勘查设计(2022年1期)2022-02-15
现代园艺(2021年23期)2021-12-01
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
今日农业(2020年19期)2020-12-14
林业调查规划(2020年3期)2020-06-03
中国科技信息(2015年2期)2015-11-16
