
哲罗鲑(Hucho taimen)卵巢发育过程中下丘脑-垂体-性腺轴(HPG轴)相关因子的动态变化
2018-12-20 01:35:56任广明佟广香张永泉徐黎明卢彤岩尹家胜
水产学杂志 2018年6期
任广明,佟广香,张永泉,徐黎明,卢彤岩,尹家胜
(中国水产科学研究院黑龙江水产研究所,黑龙江 哈尔滨 150070)
鱼类的生殖过程是在下丘脑、脑垂体、性腺组成(称下丘脑-垂体-性腺轴(hypothalamus-pituitary-gonadal axis,HPG轴)的神经与内分泌系统整体控制下进行[1]。中枢神经系统接受外界环境因子(如食物、光周期、温度、盐度、性信息素等)的刺激后,激发神经分泌细胞释放小分子神经介质传递给下丘脑,使下丘脑分泌合成促性腺激素释放激素(GnRH),促使垂体分泌促性腺激素(GtHs),即促滤泡激素(FSH)和促黄体激素(LH)[2-4]。FSH与LH在不同阶段刺激性腺产生多种性类固醇激素(雄激素、雌激素和孕激素等),具有调控配子最终成熟、促使卵母细胞或精子最后成熟、刺激排放的作用[1]。性类固醇激素除了调节卵黄和精子的发生,还能在下丘脑和垂体水平上通过正反馈刺激性未成熟鱼类GnRH和GtHs的合成与释放,启动HPG轴的功能,刺激性腺发育。可见,鱼类的性腺发育依赖于HPG轴有节奏地相互协调与制约,是神经内分泌系统调控下多种激素共同作用的生理过程。
哲罗鲑Hucho taimen是鲑形目鲑科哲罗鱼属的一种大型珍稀冷水性经济鱼类。随着全人工繁育和苗种驯化关键技术的突破,哲罗鲑养殖业迅速发展[5]。近年来,有关哲罗鲑稚鱼的生长、营养需求、胚胎与仔鱼发育、肌肉营养价值[6-9]等方面取得了一定进展,但对哲罗鲑生殖调控机制的认识仍很匮乏。随着养殖业的快速发展,如亲鱼卵巢发育不良、产卵效率和孵化率下降等问题凸显,严重制约哲罗鲑苗种生产,阻碍养殖业健康、可持续发展。神经内分泌激素协同、稳定地调控是促使雌亲鱼性腺发育成熟与产卵的关键。因此,为了深入了解HPG轴对哲罗鲑卵巢发育的调控作用,本试验测定卵巢处于不同发育期的7龄雌哲罗鲑血清中雌二醇(E2)与睾酮(T)的含量,检测下丘脑GnRH、垂体FSH与LH及肝脏卵黄蛋白原(Vitellogenin,Vtg)mRNA表达水平的变化,同时分析生殖激素基因、性腺指数(GSI)和性类固醇激素的相关性关系,以期为揭示哲罗鲑生殖调控机制提供新的思路和基础资料。
1 材料与方法
1.1 材料
试验用哲罗鲑亲鱼来自黑龙江水产研究所渤海冷水鱼试验站,挑选体质健壮、规格和体质量基本一致的同一批全人工培育的7龄雌哲罗鲑,全长70~80cm,体质量 4 500~4 700g。
1.2 采样
定期选取卵巢处于不同发育期的7龄雌性哲罗鲑,以MS-222麻醉(300mg/L)后尾部采血,血液4℃静置后于2 000r/min离心30min,收集血清于-20℃保存,用于测定血清中E2与T含量。试验鱼逐尾采血后,分别采集肝脏与性腺组织,将采集到的性腺组织快速称重用于计算GSI[100%×性腺质量/(体质量-性腺质量)]。打开试验鱼头颅,采集位于间脑后侧丘脑下方的下丘脑及丘脑末端垂体窝内垂体组织。将采集到的肝脏、性腺、下丘脑及垂体组织置于液氮中冷冻后贮存于-80℃低温冰箱中备用。
1.3 血清中E2与T含量的测定
严格按照上海酶联生物科技有限公司试剂盒的说明书,采用酶联免疫吸附法(ELISA)测定血清样品中E2和T的含量。根据标准曲线计算血清样品中激素的含量,每个处理设置3个平行。
1.4 基因的实时荧光定量PCR检测
参照 Trizol Reagent(Invitrogen,USA)说明书的方法提取下丘脑、垂体与肝脏组织RNA。采用多功能酶标仪检测RNA的浓度与纯度;利用1%琼脂糖凝胶电泳检测RNA的完整性。利用OneStepSYBRPrimeScriptTMPLUS RT-PCR Kit(TaKaRa,Japan)试剂盒测定基因表达水平变化。具体过程严格按照试剂盒说明书进行操作。根据GenBank上GnRH(大西洋鲑,Salmo salar)、LH(虹鳟,Oncorhynchus mykiss)、FSH(大西洋鲑,Salmo salar)、Vtg(大西洋鲑,Salmo salar)的基因序列,利用Premier 5.0设计荧光定量PCR特异引物,引物大小为100~150bp,引物序列见表1。以管家基因β-actin(亚东鲑,Salmo trutta fario)作为内标基因,用比较 CT 法(ΔΔCT)分析数据,取3次重复的平均值。
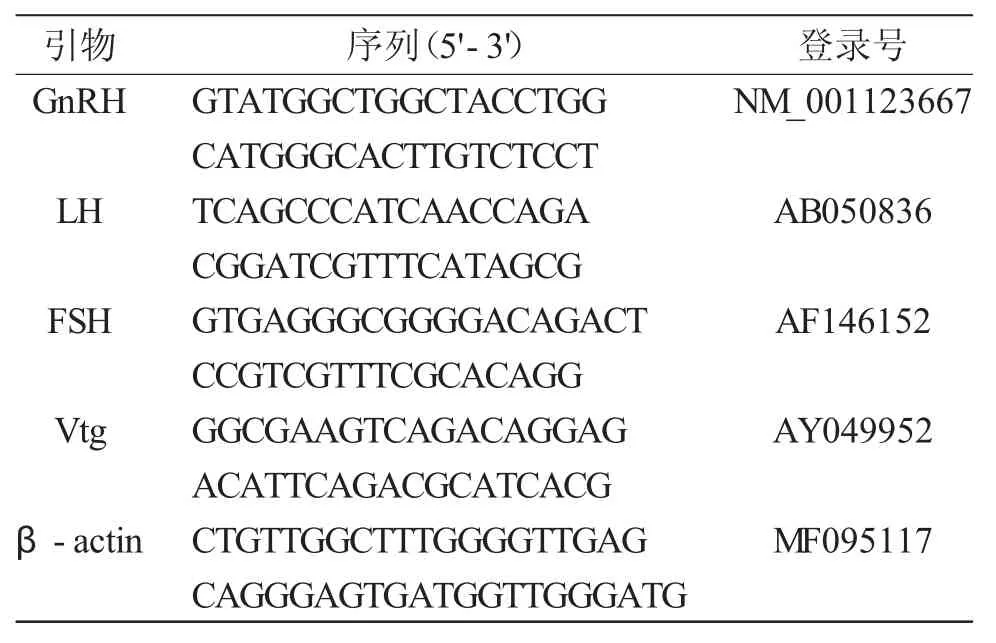
表1 哲罗鲑生殖激素基因荧光定量引物Tab.1 Primers for quantitative real-time PCR analysis of reproductive hormones in taimen
1.5 数据分析
GSI与性类固醇激素含量等数据用平均值±标准差(Mean±SD)表示,对GSI、血清性类固醇激素、相关激素mRNA表达水平分别进行单因素方差分析(One-way ANOVA)。用SPSS19.0软件对各指标进行相关性分析,P<0.05差异显著。
2 结果与分析
2.1 卵巢发育分期与GSI
以组织切片中观察到的占优势的卵细胞的发育期作为性腺的发育分期[10]。据此将采集到的哲罗鲑卵巢发育过程分为:Ⅱ期、Ⅲ期、Ⅳ期早期、Ⅳ期中期、Ⅳ期晚期、Ⅴ期早期、Ⅴ期中期。哲罗鲑不同发育期的卵巢GSI指数变化见图1。由图1可知,不同时期内卵巢GSI指数变化差异显著(P<0.05)。Ⅱ期、Ⅲ期和Ⅳ期中期卵巢的GSI相对较低(<5),Ⅳ期末期和Ⅴ期早期卵巢GSI明显上升至(7.91±1.64)%,至Ⅴ期中期卵巢时达(8.43±1.56)%。
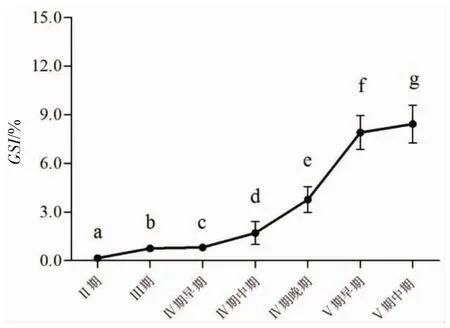
图1 哲罗鲑不同发育期卵巢的性腺指数Fig.1 Gonado-somatic index of ovary at different developmental stages in taimen
2.2 下丘脑GnRH mRNA在卵巢不同发育时期的表达
由图2-A可知,哲罗鲑产卵后(Ⅱ期卵巢)下丘脑GnRH mRNA的表达水平仍较高,随后(Ⅲ期)其表达量显著下降(P<0.05);GnRH mRNA在卵巢发育至Ⅳ期时,其表达水平呈现先升高(Ⅳ期早期)后降低趋势(Ⅳ期中期、Ⅳ期晚期)(P<0.05);卵巢发育至Ⅴ期中时,GnRH mRNA表达水平急剧升高,显著高于各阶段(P<0.05)。
2.3 垂体FSH与LH mRNA在卵巢不同发育时期的表达
Ⅱ期卵巢时,垂体FSH mRNA的表达量较低;随着卵巢发育至Ⅲ期,FSH mRNA表达量达到较高水平,随后显著下降(P<0.05)(图2-B)。卵巢发育到Ⅳ期中期,FSH mRNA表达水平再一次缓慢升高,在Ⅴ期时达峰值,随后显著下降至Ⅳ早期表达水平。由图2-C可知,在卵巢发育过程中垂体LH mRNA表达水平呈波动变化趋势,与FSH mRNA变化趋势相似。产卵后LH mRNA的表达水平较低;Ⅲ期时,表达量达峰值,随后显著下降(P<0.05)。Ⅳ期时垂体LH mRNA的表达水平较低。Ⅳ晚期时,LH mRNA表达水平再一次呈现显著升高趋势,至Ⅴ早期表达水平迅速下降。
2.4 肝脏Vtg mRNA在卵巢不同发育时期的表达
如图2-D所示,Ⅱ期和Ⅲ期时,肝脏VtgmRNA表达量较低(P<0.05)。Ⅳ期中期时,肝脏VtgmRNA表达量最高,随着卵巢的不断发育,Vtg mRNA的表达显著下降至Ⅱ期和Ⅲ期卵巢水平(P<0.05),不同发育时期差异显著(P<0.05)。
2.5 血清E2与T在卵巢不同发育期的含量变化
如图3-A所示,卵巢在Ⅱ期和Ⅲ期时,哲罗鲑血清中E2含量维持在较高水平;随着卵巢的发育,血清E2的含量显著降低,Ⅳ期早期时降至(13.93±0.14)pmol/L(P<0.05);Ⅳ期中期、晚期时,血清 E2含量保持在15.21~15.43pmol/L,卵巢发育至Ⅴ期早期时血清E2含量有所下降,降至(14.38±0.14)pmol/L;进入Ⅴ期中期时,血清E2含量逐渐升高至Ⅱ期和Ⅲ期卵巢时水平。卵巢在Ⅱ期和Ⅲ期时,血清T含量维持在60pg/mL左右(图3-B),在Ⅳ期早期其含量显著下降到(53.64±1.61)pg/mL,随后T含量显著升高,在Ⅳ期中期达到最高;进入Ⅴ期血清T含量显著下降;Ⅴ期中期血清T含量有所升高,但与Ⅳ期早期差异不显著(P>0.05)。
2.6 相关性分析

图2 哲罗鲑下丘脑GnRH mRNA(A)、垂体FSH(B)与LH(C)mRNA和肝脏Vtg mRNA(D)在卵巢不同发育期的表达Fig.2 Expression of GnRH mRNA in hypothalamus(A),pituitary FSH(B)and LH(C)mRNA and liver Vtg mRNA(D)of taimen during the ovarian development

图3 卵巢处于不同发育期的哲罗鲑血清E2与T的含量变化Fig.3 Changes in serum E2(A)and T(B)levels of taimen during the ovarian development

表2 生殖相关因子与性腺指数相关性分析Tab.2 Correlation analysis between reproductive factors and GSI
哲罗鲑卵巢发育过程中,GSI与下丘脑GnRH mRNA表达水平呈显著正相关关系,而与血清T含量呈显著负相关关系(P<0.05)。血清T含量与肝脏Vtg mRNA表达水平存在极显著的正相关关系(P<0.01)。下丘脑GnRH mRNA表达水平与垂体FSH mRNA表达水平呈极显著负相关(P<0.01),而垂体FSH mRNA表达水平与LH mRNA表达水平呈显著正相关关系(P<0.05)。
3 讨论
HPG轴作为核心的神经和内分泌系统主导着鱼类性腺发育过程中整个过程。在性腺发育及配子生成前期,感觉器官把外界的刺激通过神经联系传递到下丘脑,促使下丘脑分泌细胞的合成与分泌GnRH[1]。GnRH是HPG轴的关键神经内分泌调节因子,其mRNA表达水平在性腺发育周期内不断地变化,与性腺发育呈正相关的规律。已有研究报道,GnRH mRNA表达水平在圆斑星鲽Verasper variegatus[11]、虹鳟 Oncorhynchus mykiss[12]、平鲉 Sebastes rastrelliger[13]等鱼类卵巢发育周期内存在显著差异,在排卵和产卵期间表达水平最高,产卵后GnRH mRNA表达量显著降低,这与本研究中GnRH mRNA表达水平的变化趋势相似。垂体分泌的促性腺激素为FSH与LH[2-4]。FSH在鱼类性腺发育早期阶段主要刺激性腺分泌性类固醇激素,而LH则能促使卵母细胞最终达到成熟,是决定鱼类产卵的关键调控因子[14]。在自然条件下,高表达的GnRH能够刺激垂体合成与分泌大量的FSH与LH[15]。本研究发现,雌哲罗鲑性腺发育Ⅱ期、Ⅲ期、Ⅳ期时FSH维持相对较高的表达水平,这有利于促进性腺分泌E2和T等性类固醇激素。然而,在哲罗鲑排卵前高表达的GnRH并未促使脑垂体高表达FSH与LH,这可能是由于试验用鱼是在全人工条件下饲养,雌哲罗鲑无法进行生殖洄游,脑垂体发生功能异常等原因所致。相关性分析结果显示,垂体FSH与LH具有显著的正相关关系,这说明它们协同调控卵巢发育,能明显促进卵巢的分化与成熟。
卵黄蛋白原(Vtg)是所有卵生脊椎动物卵黄蛋白的前体,是雌鱼类卵黄的主要成分。Vtg作为一种功能性物质促进卵母细胞的生长和分化,为卵细胞和胚胎提供发育所需的营养与能源物质[16]。目前已广泛研究了鱼类性腺发育周期内Vtg的变化。在沙塘鳢Odontobutis potamophila[17]、中华鲟 Acipenser sinensis[18]和鲶 Clarias batrachus[19]产卵前(Ⅴ期卵巢)肝脏/血清中Vtg的含量最高,以确保卵巢分化、胚胎和幼体发育的营养所需。本研究结果显示,雌哲罗鲑在性腺发育早期,肝脏VtgmRNA表达相对较低,随着卵巢发育至Ⅳ期中期时,Vtg mRNA表达水平则达到最高。全人工培育条件下,雌哲罗鲑卵巢发育到Ⅲ期时出现卵黄颗粒,Ⅳ期早期时逐渐增多,到Ⅳ期中-晚期卵黄颗粒体积增大并充满卵母细胞[20]。结合本研究结果可以看出,哲罗鲑卵巢中卵黄物质的积累与卵母细胞的发育呈现同步发展的关系。肝脏中VtgmRNA的表达,促使卵黄物质迅速积累,保证营养物质向性腺转移与积累,决定卵细胞最终的发育与成熟。可见,了解雌哲罗鲑性腺发育周期内肝脏VtgmRNA表达的变化规律将为亲鱼生殖阶段营养供给提供重要的理论依据与参考。
E2与T由卵巢分泌,参与配子的发育、最终成熟和配子排放过程,是鱼类性腺成熟的关键调控因子[1]。研究发现,在斜带石斑鱼Epinephelus coioides[21]、西伯利亚鲟Acipenser baeri[22]、圆斑星鲽Verasper variegatus[23]的血浆中性类固醇激素(E2与T)含量在排卵前显著升高,排卵后迅速下降。本研究中,雌哲罗鲑在Ⅱ期卵巢时血清E2与T含量相对较低,说明亲鱼正处于恢复生长和营养积累的阶段。Ⅳ期卵巢是卵子卵黄发生的关键时期,大量合成Vtg,随后T将Vtg运输到卵细胞内[1],这说明E2与T参与了配子卵黄发生的调控过程[24,25]。本研究中,随着哲罗鲑卵母细胞中卵黄发生基本完成,血清中E2与T含量降低,当亲鱼卵巢发育至Ⅴ期时,血清中E2与T含量明显增加,这在一定程度上说明E2与T在亲鱼产卵前起正向调节性腺发育的功能。
综上所述,GnRH、FSH与LH以协同的方式共同参与了哲罗鲑卵巢发育的过程,其调控作用与Vtg、E2与T的合成与分泌紧密相关。了解哲罗鲑卵巢发育过程中性激素因子的变化规律,将有助于系统、科学地开展哲罗鲑人工繁育与健康养殖。然而,在全人工饲养条件下,哲罗鲑性腺发育受到了诸多因素的影响,HPG轴的调控机制还需结合环境因子、人为干预等因素进行深入的探讨与研究。
猜你喜欢
文萃报·周五版(2022年24期)2022-06-21 20:55:40
昆明医科大学学报(2020年12期)2021-01-26 00:44:02
中老年健康(2017年9期)2017-12-13 07:16:39
中成药(2017年6期)2017-06-13 07:30:34
海南医学(2016年8期)2016-06-08 05:43:00
中国民族医药杂志(2016年7期)2016-05-09 07:49:19
磁共振成像(2015年9期)2015-12-26 07:20:34
中学科技(2014年11期)2014-12-25 07:27:11
食品工业科技(2014年15期)2014-03-11 18:17:46
微创泌尿外科杂志(2014年4期)2014-02-28 17:28:44
