
鄱阳湖越冬雁类食源植被适宜取食时间窗口
2018-12-19 10:44孟竹剑夏少霞于秀波饶滴滴金斌松
生态学报 2018年21期
孟竹剑,夏少霞,于秀波,,*,饶滴滴,金斌松,
1 南昌大学生命科学研究院流域生态研究所和生命科学学院,南昌 330031 2 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室,北京 100101 3 江西师范大学地理与环境学院,南昌 330022 4 南昌大学鄱阳湖环境与资源利用教育部重点实验室,南昌 330031 5 江西鄱阳湖湿地生态系统国家定位观测研究站,南昌 330038 6 中国科学院大学,北京 100049
长江中下游湿地是世界自然基金会确定的生物多样性保护的热点区域之一[1],也是东亚-澳大利西亚迁飞路线上重要的越冬地,每年吸引约数十万只候鸟来此越冬[2]。其中,雁鸭类是越冬水鸟中的优势类群[3],主要以白额雁(Anseralbifrons),小白额雁(A.erythropus),豆雁(A.fabalis)和灰雁(A.anser)为主[4- 5]。雁类到达越冬地时,一方面,由于经过长距离的迁徙消耗了大量能量,处于能量匮乏期,另一方面需要为春季迁徙储存充足的能量。因此,越冬期的能量和营养补充对雁类完成迁徙的种群保育至关重要[6]。
由于能量平衡和营养的需求,植食性水鸟对取食植被具有高度的选择性,植被高度以及营养成分含量会影响水鸟对食物资源的选择[7]。在洪泛平原湿地,受季节性水文波动的影响,植被发育与洲滩淹没和出露时间密切相关[8]。莎草科植被苔草(Carexspp)是长江中下游越冬雁类的主要食物资源[9],每年有春季和秋季两个生长期[10]。然而并不是所有生长阶段的苔草植被均能被雁类取食。研究表明,苔草的高度、纤维素和蛋白质的含量[11]以及“鲜绿”程度会影响雁类对取食斑块的选择[12]。然而,这些特定的取食特征与洲滩出露时间的关系尚缺乏可考证资料。
本研究拟建立雁类取食植被的特征和苔草生长过程及退水时间的关系,从而为雁类栖息地的划分和保护提供定量的评估指标。由于株高和生物量便于观测,且与水文变化的关系更为直观[13],同时,营养元素的含量往往与也与植被的生长阶段相关[14],因此,本研究主要选取了株高和生物量这两个生长因子作为取食植被的特征。
本研究选取位于长江中下游的鄱阳湖国家级自然保护区的常湖池作为研究区,通过原位观测试验,辅以遥感影像、数字高程模型(Digital Elevation Model,DEM)、气温等数据资源,结合野外观测和室内分析,揭示了苔草“春草”的生长过程,同时结合已发表文献数据,补充了对“秋草”生长过程的估算,量化洲滩出露时间与苔草株高之间的关系,确定了适宜雁类取食植被的时间窗口。本研究对评估栖息地质量,预测栖息地负载量和制定相关保护策略具有重要意义。
1 研究区概况
鄱阳湖位于江西省北部,是我国第一大淡水湖。湖区属于亚热带湿润季风型气候,气候温暖湿润,雨量充沛,年平均降水量为1350—2150 mm,年平均气温16.5—17.8℃。鄱阳湖国家级自然保护区,以保护珍稀候鸟和湿地生态环境为目标,1992年7月被国际重要湿地公约秘书处列为国际重要湿地。保护区辖有9个湖泊,面积22400 hm2,占全湖面积的5%,但却承载了全湖越冬候鸟总数量的70%[15- 16]。
研究区设在鄱阳湖国家级自然保护区的常湖池(图1),常湖池面积约700 hm2,是季节性的浅碟形子湖泊之一[17]。常湖池面积适中,湖水较浅,从岸边到湖心方向,水位梯度较为明显,植被以莎草类群落为主,呈明显的不规则环带状分布。周边泥滩和草洲面积相对较大,是雁类在鄱阳湖的主要分布区之一[16]。
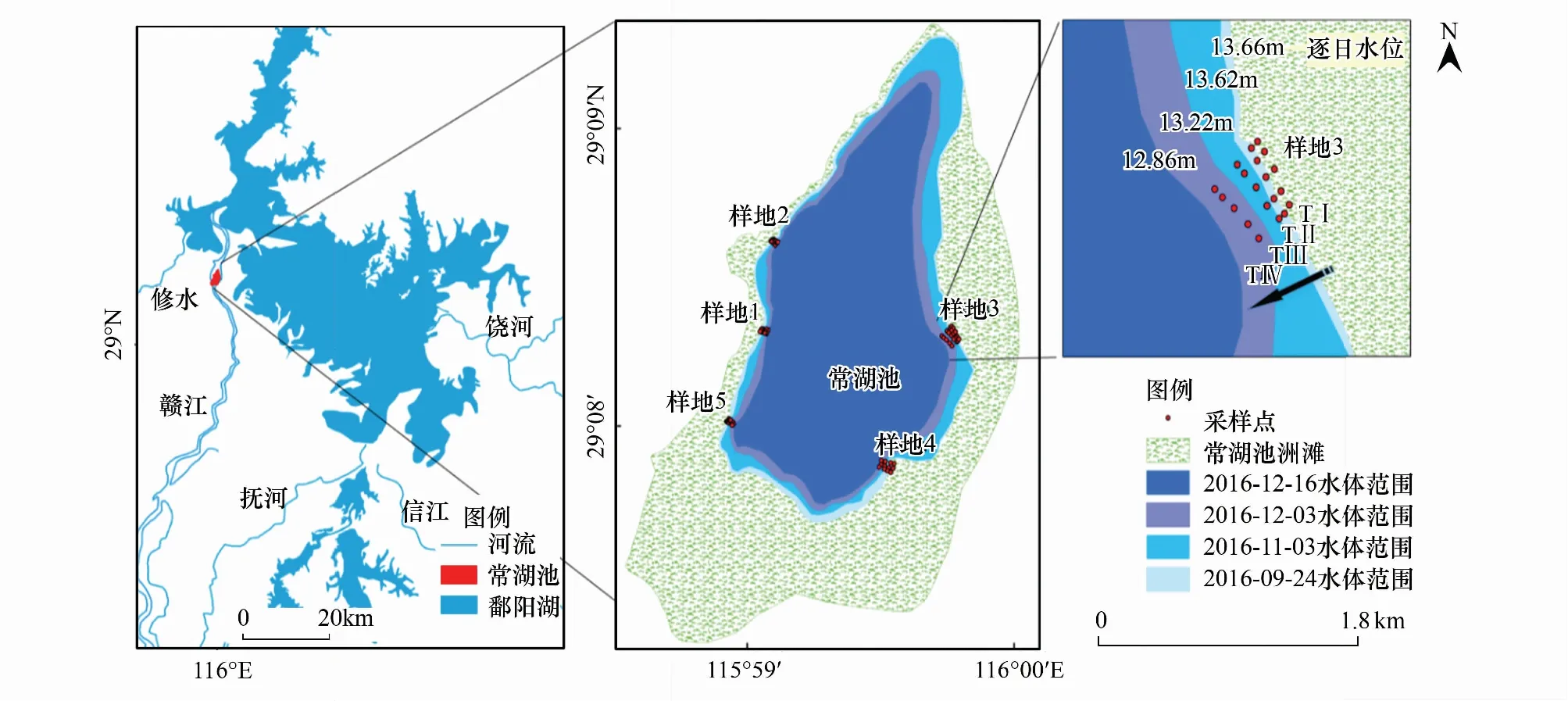
图1 研究区示意图Fig.1 Location of study area图中水体范围为遥感影像解译的结果,水位为遥感影像获取时间对应的常湖池观测水位,数据来自江西省水文局
2 数据与研究方法
2.1 样品采集与处理
根据前期的实地调查,选择越冬期雁类数量较多的区域,于“春草”生长期开始前(2017年1月18日),环湖布设5个原位监测样地,结合常湖池退水过程和数字高程模型,在各监测样地从岸边到湖心沿等水位线设立4个梯度(TⅠ—TⅣ,高梯度—低梯度),每个梯度均设有17个平行样方,共68个(图1),其中样地1、2、4、5每个梯度均为3个平行样方,样地3由于越冬期雁类数量较多,分布较为集中,每个梯度设置5个平行样方,样方规格均为1 m×1 m。为避免雁类取食和动物践踏等外部因素干扰,样方外围设置了网罩,网目规格为60 mm。
2017年3月8日—2017年5月2日,对苔草的“春季生长期”开展监测,监测时间共计52 d。采样频度及具体监测指标见表1。采样过程中,在每个实验样方内随机选择10株苔草植株,采用现场测量法测定株高。测量完成后,再在实验样方周围选择株高和盖度一致的植株,采用收割法,采集植物地上部分,取样规格为0.25 m×0.25 m,装入信封编号后称湿重,带回实验室。将采集的样品置于烘箱,60℃烘至恒重,测定并记录植株干重、含水量。

表1 采样时间及监测指标
“√”表示采样得到的监测指标
2.2 洲滩出露时间确定
本研究采用欧空局发布的Sentinel- 2遥感数据(http://www.gdal.org/frmt_sentinel2.html)监测判定洲滩出露时间。该卫星能够提供高空间分辨率(10、20 m和60 m)的多光谱数据(13个波段),其中,红外区域设置3个波段能更好的反映植被变化情况。与Landsat 数据相比,Sentinel- 2 遥感数据空间分辨率高、重访周期短,能满足在水体、植被信息提取及退水时间监测方面的需求[18]。本研究选取了研究区范围内云量较少、质量较好的8景影像,时间从2016年9月25日—2017年4月22日,根据鄱阳湖水文节律,退水期为9月—11月,影像覆盖了退水期至春草生长结束期。在e Cognition 8.7中分别对8景进行水边界提取,将监测样点与数字高程模型和提取的水体边界进行叠加,如样点被水体覆盖则未出露,反之,则出露。由此,得到常湖池监测样点的出露时间从高梯度到低梯度分别为2016年9月24日,2016年11月3日,2016年12月3日和2016年12月16日。
2.3 苔草有效生长时间确定

图2 逐日平均温度变化Fig.2 The changing of daily average temperature
研究发现,苔草生长对气温比较敏感,日平均温度低于10℃时,处于停滞萌发和生长状态[19]。因此,使用该区域的逐日气温数据(数据来源于江西师范大学地理与环境学院),判别自洲滩出露后气温大于10℃的天数作为苔草的有效生长时间。整个越冬期,自11月20日开始(图2),温度降至10℃以下,低温状态从2016年11月底断断续续地持续到3月初,持续时间为100 d。此外,研究表明,苔草种子在(20±5)℃条件下,萌发(以芽尖伸出土面为发芽标准)时间需要5 d左右[20],因此,在有效生长时间中,应将低温持续天数和萌发时间扣除。
2.4 雁类取食植被生长特征的确定
2017年越冬期,多次在鄱阳湖大湖池、常湖池、白沙湖等子湖泊开展雁类栖息地及取食食物资源特征调查。根据实地观测、雁类脚印、新鲜粪便等确定雁类觅食地,根据取食苔草叶片的痕迹判定是否为新鲜取食,并对该觅食地取食、未取食的植株株高进行现场测量,并记录。共计获得取食、未取食株高数据540个,选择被取食苔草的最高值和最低值作为雁类取食范围,即7.5—18.7 cm(图3)。
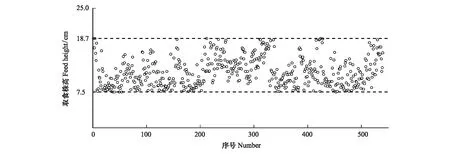
图3 基于调查数据的雁类取食苔草株高分布图Fig.3 Height of Carex spp distribution after geese feed based on survey data
2.5 植被生长过程模拟及时间窗口确定
植被生长过程中,其植株的高度、生物量及营养含量(如蛋白质)等的变化曲线都近似于“S”形,即符合Logistic方程,因此该生态学模型被广泛用于模拟植物生物量及生长过程[21]。本研究采用Logistic方程分别对苔草的株高和地上生物量的季节动态进行拟合。
(1)
式中,y是植株的株高或地上生物量;t是有效生长天数;k为环境容纳量最终可达的最大值;a,b为待定系数,e为自然对数底。
在模型参数的求算中,K值的确定是最为重要的。目前常见的方法有目测法、三点法、四点法、均值法等[21]。研究表明,三点法、四点法、均值法均能达到较高的拟合精度,其中以四点法最优[22]。四点法即用实测序列中的4个数据点来估计K值。四点法估计K值的公式为:
(2)
t1+t4=t2+t3。式中(t1,N1)、(t4,N4)分别为实测数据序列的始点、终点,(t2,N2)、(t3,N3)则为中间两点。
由于本实验缺少了对“秋草”生长阶段的观测,此阶段苔草生长过程采用已发表文献的数据[18],得到0—61 d的生长过程曲线(图4),该实验设计温度在(20±5)℃条件下,淹水2 cm,实验条件符合苔草秋季生长期的实际状况。拟合方程为:
f(t)=28.46/(1+exp(1.779-0.1067×t))R2=0.993
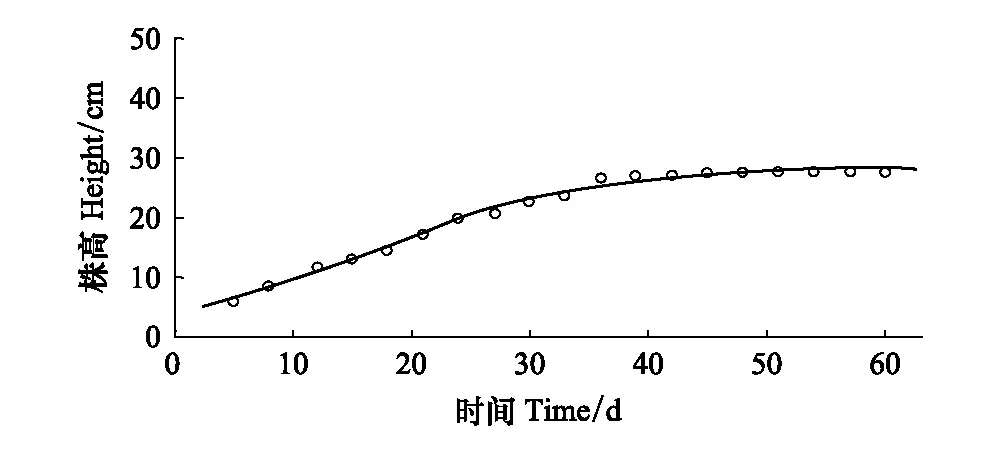
图4 秋季生长期苔草株高随时间的变化 Fig.4 Temporal variation of plant height of Carex spp in autumn
将苔草“春季生长期”株高和地上生物量指标在Excel 2013中进行数据预处理,在Matlab中用双因素方差分析,分析4个水分梯度对苔草生长过程影响的差异和不同样地间苔草生长过程的差异性,然后在Matlab中分别对4个高程梯度的生长曲线进行拟合。
根据拟合方程,结合雁类取食苔草的株高范围、洲滩出露时间和气温确定的有效生长天数,对生长模型方程进行求解,即可得到雁类取食的时间窗口。
计算公式为:T= [Tmin,Tmax]
(3)
式中,Tmin=tmin-t1-t2,,Tmax=tmax-t1-t2
(4)
(5)
式中,k和a,b值同公式1,7.5和18.7分别对应被取食苔草的最高值和最低值,t1为从出露开始,温度低于10℃的天数(100 d),t2为萌发所需要的天数,本研究取5 d。
3 结果与分析
3.1 苔草株高和地上生物量动态变化
2017年3月份,TⅠ、TⅡ、TⅢ、TⅣ苔草的初始测量值平均株高为((11.07±3.2) cm,(8.77±2.54) cm,(8.15±2.7) cm,(8.99±2.87) cm),地上生物量为((92.49±48.85) g/m2,(97.70±36.27) g/m2,(116.07±33.1) g/m2,(102.31±29.25) g/m2)。实验期间,苔草的平均株高和地上生物量逐渐增加。并于5月达到最大值,此时TⅠ、TⅡ、TⅢ、TⅣ平均株高分别为((60.95±5.73) cm,(58.63±6.48) cm,(52.74±5.81) cm,(53.19±6.84) cm),地上生物量分别为((598.8±141.52) g/m2,(537.8±147.97) g/m2,(500.2±112.9) g/m2,(428.4±76.1) g/m2)。各梯度苔草平均株高(F=4.31,P<0.05)和地上生物量(F=9.28,P<0.05)差异性显著,样地之间苔草平均株高(F=6.4,P<0.05)和地上生物量(F=10.62,P<0.05)也呈现显著差异。当苔草生长达到最大值时,各梯度株高表现为TⅠ>TⅡ>TⅣ>TⅢ,地上生物量表现为TⅠ>TⅡ>TⅢ>TⅣ(图5)。苔草的株高和地上生物量符合Logistic增长模型,且拟合效果较好,相关系数值均在0.92以上(表2)。
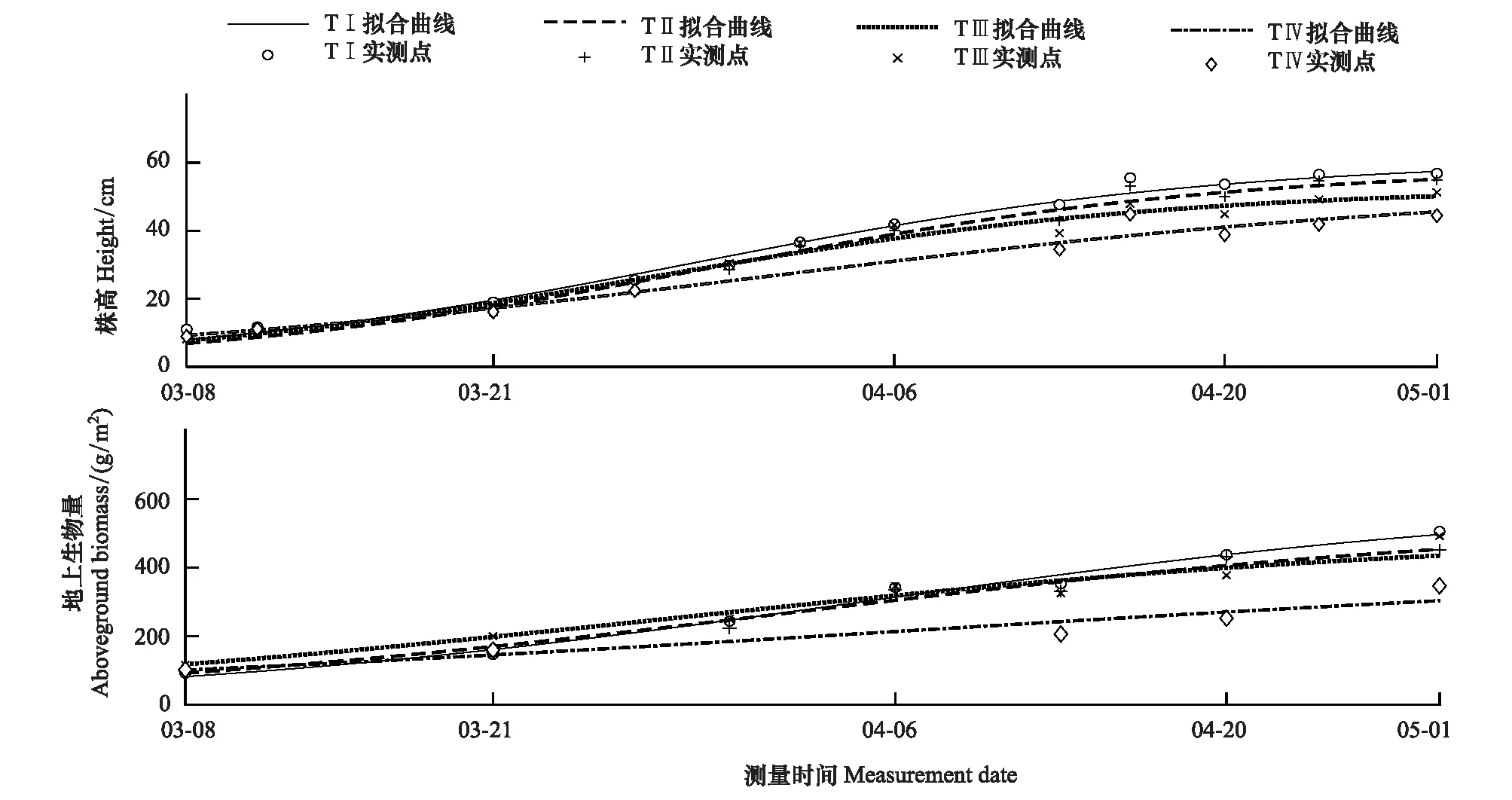
图5 苔草株高和地上生物量春季生长过程曲线Fig.5 The growth curve of height and aboveground biomass of Carex spp in spring
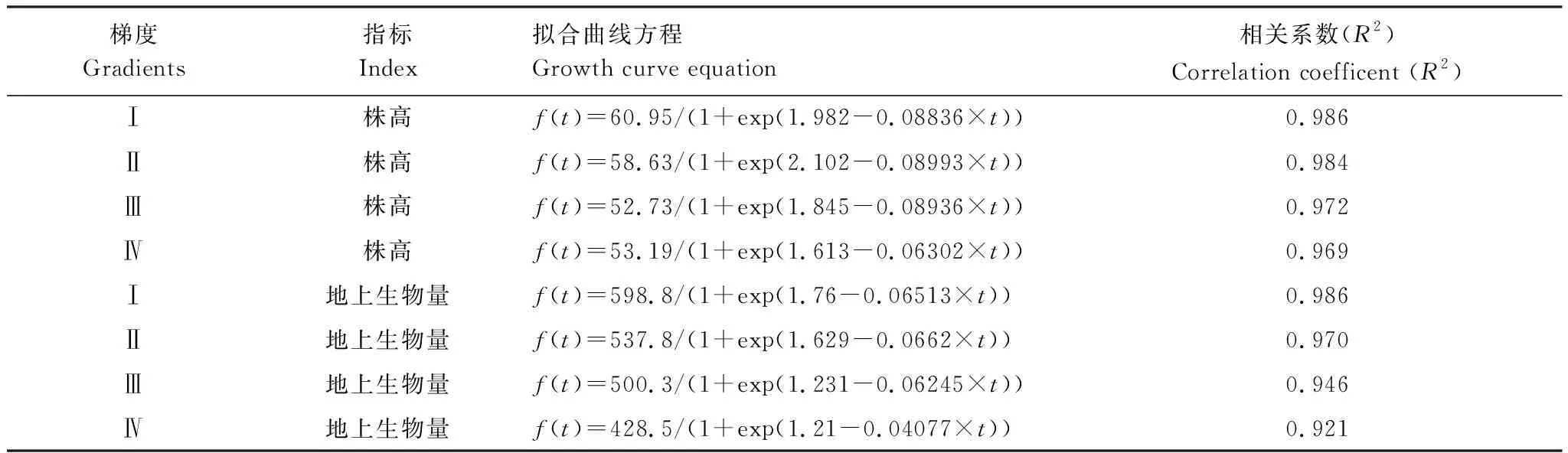
梯度Gradients指标Index拟合曲线方程Growth curve equation相关系数(R2)Correlation coefficent (R2)Ⅰ株高f(t)=60.95/(1+exp(1.982-0.08836×t))0.986Ⅱ株高f(t)=58.63/(1+exp(2.102-0.08993×t))0.984Ⅲ株高f(t)=52.73/(1+exp(1.845-0.08936×t))0.972Ⅳ株高f(t)=53.19/(1+exp(1.613-0.06302×t))0.969Ⅰ地上生物量f(t)=598.8/(1+exp(1.76-0.06513×t))0.986Ⅱ地上生物量f(t)=537.8/(1+exp(1.629-0.0662×t))0.970Ⅲ地上生物量f(t)=500.3/(1+exp(1.231-0.06245×t))0.946Ⅳ地上生物量f(t)=428.5/(1+exp(1.21-0.04077×t))0.921
3.2 株高与地上生物量之间的关系
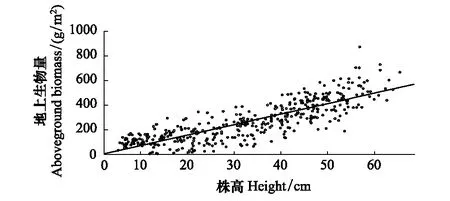
图6 苔草的株高和地上生物量的关系 Fig.6 The fitting curve of Carex spp relationship between height growth and aboveground biomass
对苔草的株高和地上生物量进行拟合分析(图6),表明株高和地上生物量之间存在显著正相关(R2=0.70,P<0.01)。
苔草株高生长与地上生物量关系及拟合方程为f(x) =8.529x-14.48。
3.3 雁类取食时间窗口的确定
根据确定的各梯度的出露时间,减去该阶段停滞生长时间(100 d)和苔草从出露到萌发的时间(5 d),即为正常状态下苔草的有效生长天数。由此得到,秋季生长期,TⅠ、TⅡ在出露天数达到12 d时适宜雁类取食,出露时间超过28 d后,不适宜雁类取食,TⅢ、TⅣ在秋季生长期出露后处于低温状态,无法萌发。在春季生长期,TⅠ、TⅡ、TⅢ、TⅣ在出露天数分别达到169—182、131—144、99—112、83—102 d时适宜雁类取食(图7和表3)。
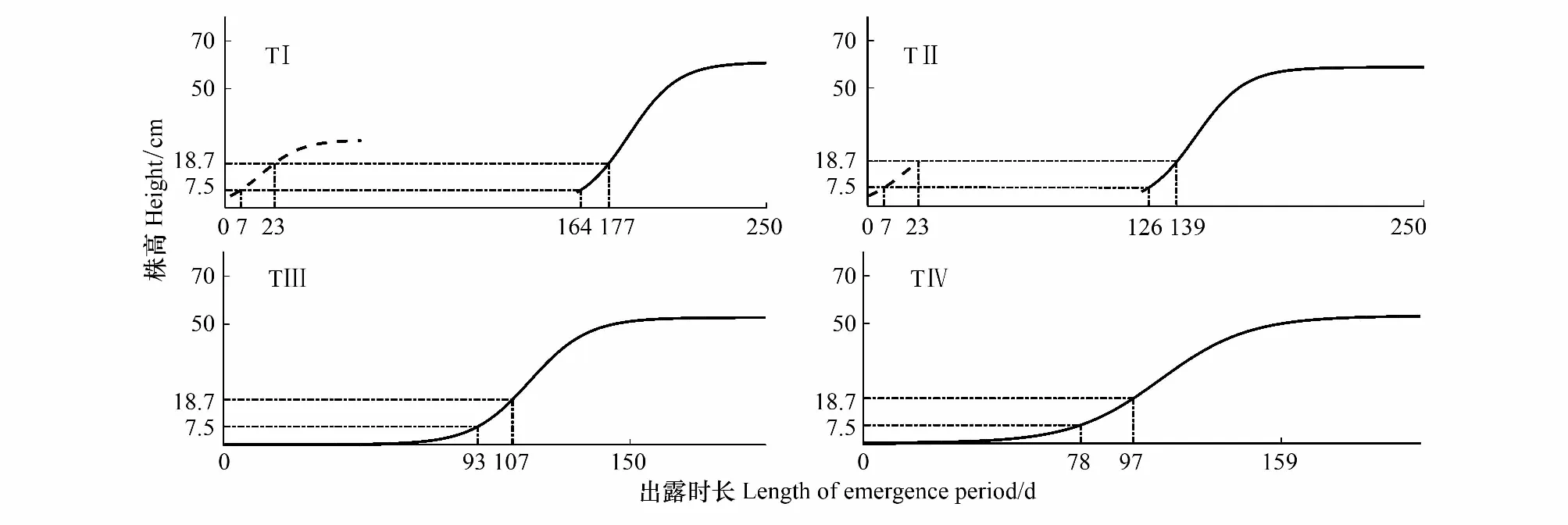
图7 越冬雁类适宜取食草洲的时间窗口Fig.7 The suitable time window for overwintering geese to feed on

梯度Gradients生长曲线方程Growth curve equation生长时间Growth time/d停滞生长时间Stagnant growth time/d萌发时间Germination time/d有效生长时间Effective growth time/dⅠf(t)=28.46/(1+exp(1.779-0.1067×t))t<612181005113低于有效温度,停滞生长61≤t≤161f(t)=60.95/(1+exp(16.47-0.08836×t))t>161Ⅱf(t)=28.46/(1+exp(1.779-0.1067×t))t<17175100578低于有效温度,停滞生长17≤t≤118f(t)=58.63/(1+exp(13.25-0.08993×t))t>118Ⅲf(t)=52.73/(1+exp(10.59-0.09374×t))12761561Ⅳf(t)=53.19/(1+exp(6.718-0.06302×t))12155561
4 讨论
4.1 洲滩出露时间对苔草生长过程的影响机制
对雁类而言,植株高度、纤维素和蛋白质的含量[11]以及“鲜绿”程度是影响其取食的重要因素[12]。其中,植株高度是影响植食性鸟类的啄食速率[23],限制其食物的可利用性的关键指标[24]。此外,随着植被高度及生物量逐渐增加,纤维和木质素含量也随之增加,而可消化能量的含量反而降低[25]。而植被的氮浓度[26]、易消化有机物及磷素[27]等与植被生物量等生长特征也密切相关。因此,株高和生物量指标不仅能直观反映植物生长状况,也能一定程度上反映植被营养状况,指示植食性生物的食物资源偏好特征。
研究发现苔草株高和地上生物量的生长均为Logistic增长模型,拟合效果较好,并且二者呈显著正相关,而利用Logistic方程拟合苔草株高的生长曲线优于地上生物量的拟合效果(图5)。这可能与生长过程中一些器官枯死凋落、营养物质溶失及向地下根系转移,致使地上生物量逐渐下降有关[28]。此外,野外原位实验中,自然环境因子难以控制,也会对植株生长过程造成一定影响。
影响湿地植被生长最主要的因子为温度和水分条件。温度影响植被的萌发和生长速率[29]而温度低于植物生长所需有效积温时,植物停滞生长[30- 32]。而水位梯度也是导致植株生长曲线差异的原因。水深及淹水时长不仅影响植被生物量的形成[33],还会影响地上和地下生物量的分配[34- 35]。而在放水过程中,不同高程梯度样地的地下水位从高梯度到低梯度的逐渐下降,这种变化导致土壤等生长环境及植物的生物量分配产生差异[36]。不同高程洲滩由于出露时间的差异,导致水分梯度以及萌发和生长阶段的有效积温产生差异,从而对生长曲线产生影响。本研究在生长曲线的拟合中,考虑了温度和退水时间对生长的影响,能更准确刻画鄱阳湖湿生植被生长过程。此外,不同样地之间苔草株高和地上生物量之间的差异,可能是由于土壤基质等因素造成的。研究期间,赣江上游降雨量突增,导致水位升高,江水倒灌,使得TⅢ、TⅣ被水淹没。退水后,部分淹水植株叶缘发黄受损,致使株高、地上生物量降低,对其生长过程曲线有一定影响。
4.2 退水时间的差异与雁类对食物资源的利用

图8 雁类取食痕迹(拍摄于2016年12月8日) Fig.8 The feeding trail for overwintering geese (Picture was taken in 8th December, 2016)
本实验中,TⅠ样地最早出露,时间为9月下旬,按照生长曲线确定的窗口期,在雁类到达鄱阳湖(10月上旬)时,该样地苔草处于适宜取食的阶段(图7和表3)。而TⅡ在11月3日出露萌发,并且在低温到来前,该梯度苔草仍处于适宜雁类取食区间范围内,由于该梯度面积较大,因此,在整个越冬期,TⅡ都作为雁类主要的潜在觅食地。调查发现,低温到来之后,该样地雁类的取食痕迹特别明显,几乎将整片草洲尽数啃食(图8)。同时,根据本研究,若推迟退水时间达到20 d,即本研究中的TⅢ、TⅣ,尽管在12月相继出露,然而,此时受低温的限制,苔草种子无法萌发,仍埋在土壤中,无法满足雁类在越冬季(12月至次年2月)的取食需求。本研究与鄱阳湖湿地植物的生长发育过程研究结果一致[37-38],表明退水时间的差异对雁类可取食食物资源有影响。相关研究也认为,过早或推迟退水时间均会影响苔草生长,无法满足雁类的取食需求,导致食物资源短缺[10]。而对雁类越冬行为的观测表明,其在秋季积累能量,冬季消耗能量[39],当觅食地的食物供给无法满足其需求时,雁类会转向觅食低密度、低植株高度和低生物量的植被或其他食源植物[40- 41]。因此,合理调控子湖泊放水过程,确保植被生长区间与雁类迁徙节律的匹配性,将有利于保护雁类种群维持[42]。本研究对雁类取食植被株高和地上生物量因子展开研究,食源植被的营养供给也是植食性雁类选择栖息地的重要因素,需要进一步对雁类栖息地进行更深入的研究,为雁类和其他水鸟的保护提高基础研究支撑。
5 结论
苔草生长符合Logistic增长曲线,且株高因子的拟合效果更好。株高与地上生物量显著性相关,这表明采用株高可用于预测苔草地上生物量动态。不同梯度苔草生长曲线的差异由水位梯度和气温因素共同决定的,结合雁类取食的特征,秋季生长期苔草适宜雁类取食的时间窗口为12—28 d;而春季生长期,时间窗口则为83—182 d。这意味着在9月下旬出露的洲滩,将于10月中下旬达到雁类取食的时间窗口,这些洲滩苔草的生长节律与雁类到达的时间匹配。而推迟退水时间,将由于低温限制,导致苔草的有效生长积温减少,延长苔草到达适宜雁类取食窗口的时间。而从11月中旬开始,气温趋于10℃以下,因此,推迟退水时间20 d,将使苔草到达适宜雁类取食时间窗口的时间延长3个月以上,难以满足12月、1月雁类到达觅食高峰期的需求。本研究对鄱阳湖湿地水文调控和湿地资源管理具有重要意义。
致谢:江西师范大学提供数据支持,刘宇、张欢、张广帅、段后浪修改论文,特此致谢。
猜你喜欢
农业与技术(2022年24期)2023-01-05
水利水电快报(2022年7期)2022-07-18
草地学报(2021年6期)2021-07-05
现代出版(2019年6期)2020-01-14
农村百事通(2019年17期)2019-10-08
湖泊科学(2017年4期)2017-07-31
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
老友(2017年4期)2017-02-09
