
基于流式细胞术的旱柳染色体倍性与基因组大小测定
2018-12-19 08:50:44马祥建李玉娟
江苏农业科学 2018年22期
张 健, 王 莹, 马祥建, 谈 峰, 李 敏, 李玉娟
(江苏沿江地区农业科学研究所,江苏南通 226541)
柳树是杨柳科(Salicaceae)柳属(Salix)树种,为落叶乔木或灌木,主要分布在北半球温带及寒带地区,在北半球热带、亚热带地区及南半球也有少量分布。全球有500多个柳树品种,其中亚洲是柳树的起源中心,仅我国就有275种[1]。柳树树种间存在丰富的染色体倍性差异,包括二倍体(2n=2x=38)、四倍体(2n=4x=76)、六倍体(2n=6x=114)甚至十二倍体(2n=12x=228)[2],其中灌木柳主要为二倍体,乔木柳多为异源多倍体[1,3]。
旱柳(Salixmatsudana)是柳属最重要的树种之一,主要为乔木柳[4],原产我国东北部,又被称为中国柳,以其高适应性特别是抗逆性已被引种推广到澳大利亚、欧洲与北美洲等地区[5],是重要的能源、造林和绿化树种[6]。旱柳在土壤重金属修复、盐碱地绿化、生物燃料等方面发挥着重要作用[7-8],其中海柳1号、9901柳等品种已逐渐成为我国沿海林业开发中潜力巨大的战略树种[9-10]。目前,科研人员已开展了旱柳生理指标如超氧化物歧化酶(superoxide dismutase,简称SOD)活性、过氧化氢酶(peroxidase,简称POD)活性、丙二醛(malondialdehyde,简称MDA)含量在盐胁迫下的表现[9]以及转录组[11]、比较蛋白组[5]及miRNA[12]等方面的研究,但旱柳的染色体倍性与基因组大小尚未见报道,这限制了旱柳分子育种的发展。
流式细胞术(flow cytometry,简称FCM)是一种在液流系统中快速测定单个细胞或细胞器的生物学性质,并把特定的细胞或细胞器从群体中加以分类收集的技术,其特点是通过快速测定库尔特电阻、荧光、光散射和光吸收来定量测定细胞染色体倍性、DNA含量、细胞体积、蛋白质含量等许多重要参数[13]。流式细胞术已在山药、芭蕉等植物的染色体倍性检测与基因组大小测定中发挥了重要的作用[14-15]。
本研究运用流式细胞术对旱柳不同品种的染色体倍性与基因组大小进行测定,以期为旱柳全基因组测序提供参考。
1 材料与方法
1.1 试验材料
旱柳品种选用江苏省地方旱柳品种沿江柳、江苏沿江地区农业科学研究所育成的旱柳品种海柳1号、山东省林业科学研究院育成的旱柳品种9901柳。旱柳染色体倍性测定以东北地方灌木柳品种杞柳(已知为二倍体)为对照,基因组大小测定以基因组大小已知的水稻淮稻5号(江苏徐淮地区淮阴农业科学研究所育成)和大豆品种通豆7号(江苏沿江地区农业科学研究所育成)为对照。
1.2 培养条件
染色体倍性测定时,分别取杞柳与旱柳各品种的茎段扦插条(粗2~3 cm,长8~10 cm),放入含1/3清水透明杯中并置于光照培养箱中培养,培养条件:光暗周期为12 h—12 h,光照度为3 300 lx,培养温度为25 ℃。
基因组大小测定时,取水稻品种淮稻5号、大豆品种通豆7号的籽粒,用培养皿加水后放入光照培箱内培养;同时取旱柳3个品种茎段,培养方式与上述一致。
1.3 染色体倍性及基因组大小测定
上述材料培养时间均为20 d,培养后取新鲜嫩叶,采用FACSCalibur流式细胞仪(美国BD公司)进行染色体倍性和基因组大小测定,并用CellQuest软件(美国BD公司)获取数据,用ModFit软件(Yerity Software House公司)分析结果。
单细胞悬液:取1 g植株新鲜叶片,在2 mL细胞裂解液中用锋利的刀片切碎,用200目尼龙网过滤,用试管收集滤液,经1 000 r/min离心5 min后,弃去上清液,收集沉淀细胞核。
混合液:先用碘化丙啶(propidium iodide,简称PI)染液对细胞核重悬,将细胞核悬液DNA进行荧光标记,置于暗处染色 20 min。再将几个品种的染液进行混合形成混合液。
用已知染色体倍性或基因组大小的材料作为对照,将对照材料横坐标固定,保持各个参数一致后对待测样品逐一检测,根据峰的位置并参考对照样品的大小计算待测样的倍性与基因组大小。
2 结果与分析
2.1 旱柳染色体倍性分析
用流式细胞仪测定杞柳(二倍体)的DNA含量时将其设定为50附近,以二倍体杞柳的DNA含量为对照,将其他待测旱柳品种的DNA含量与其进行比较,得到所测样品的相对DNA含量。从图1可以看出,杞柳在50左右有1个主峰(图1-a),海柳1号、9901柳和沿江柳3个旱柳品种均在100左右出现了主峰,代表其倍性约为杞柳的2倍,即3个旱柳品种均为四倍体。
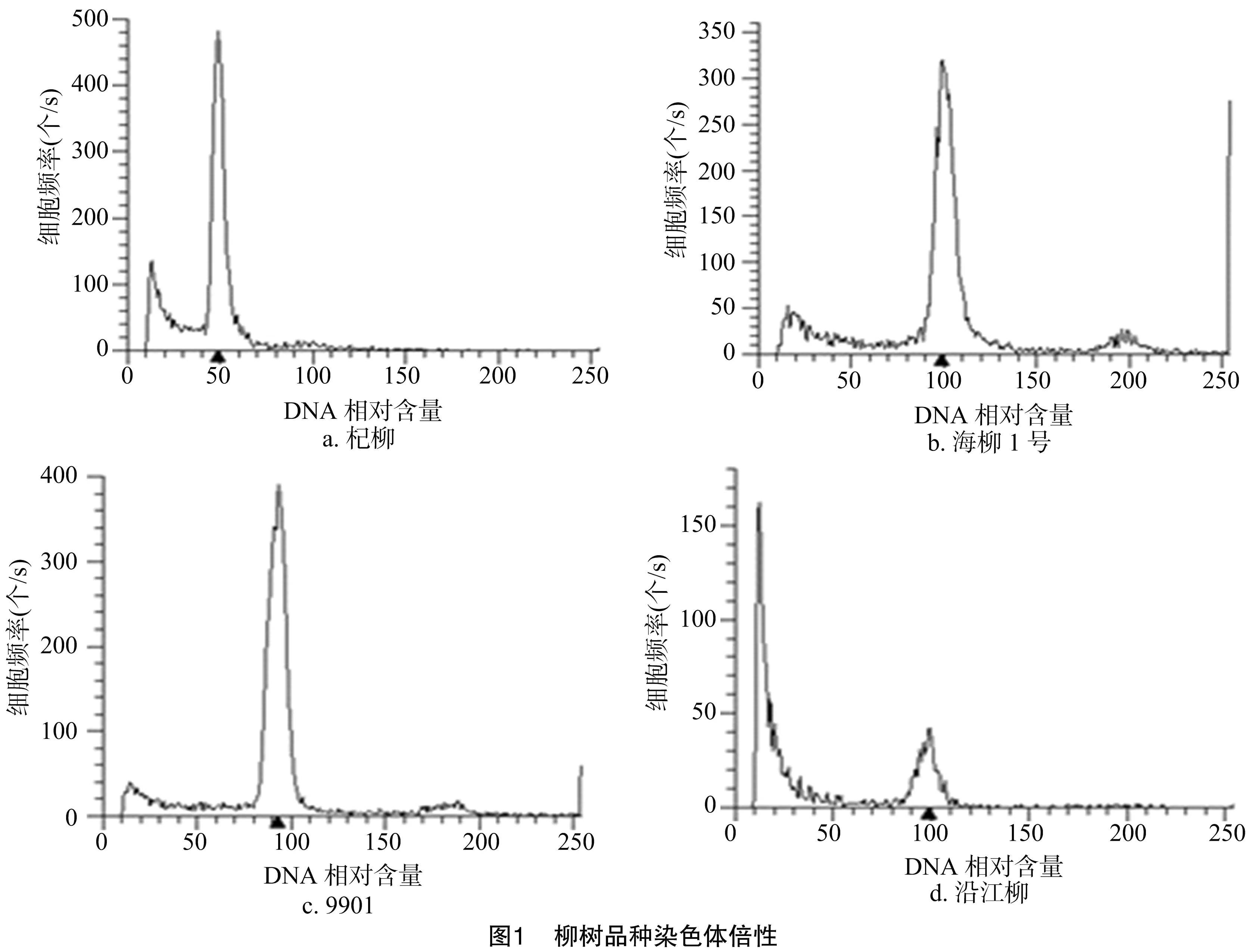
2.2 旱柳基因组大小预测
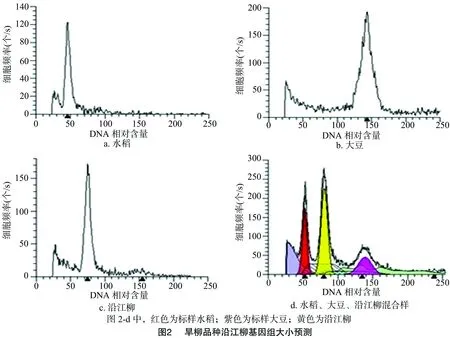
2.2.1 水稻、大豆、沿江柳基因组比较 用流式细胞仪测试水稻细胞的单细胞悬液,已知水稻基因组大小约为 440 Mb[16],所得的DNA含量定为44左右,并以此为对照,测定大豆(已知其基因组大小约为1 150 Mb[17])和沿江柳的DNA含量,并将沿江柳的DNA含量分别与水稻和大豆的DNA含量进行比较,得到沿江柳的相对DNA含量,结果见图2-a、图2-b、图2-c。用流式细胞仪对沿江柳与水稻、大豆进行了混合样测定,将最小基因组的水稻DNA定量为50左右,得出图2-d的混合样图。
从图2-a、图2-b、图-2c可以看出,水稻的G1期(细胞分裂间期1)在图2-a中横坐标46.49处,大豆的G1期位置在143.56处,沿江柳的G1期位置在75.60处,从水稻推算沿江柳的基因组大小为440/46.49×75.60=715.5 Mb;从大豆推算沿江柳的基因组大小为1 150/143.56×75.60=605.6 Mb。
从图2-d可以看出,水稻G1期的位置在52.27处;大豆G1期位置在138.74处,沿江柳的基因组介于两者之间,峰值在80.44处。从水稻推算沿江柳的基因组大小为440/52.27×80.44=677.1 Mb,从大豆推算沿江柳的基因组大小为1 150/138.74×80.44=666.8 Mb。
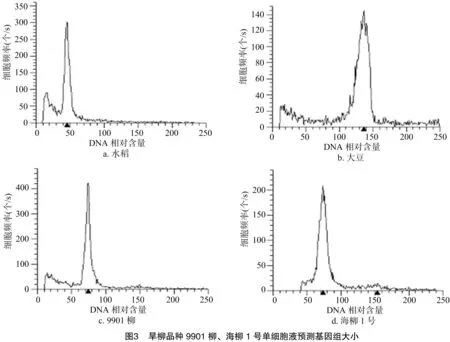
2.2.2 水稻、大豆、9901柳、海柳1号单细胞悬液比较 为了更准确地测定出旱柳基因组大小,笔者重复测定了水稻、大豆的单细胞悬液,并将9901柳、海柳1号这2个旱柳品种与其进行比较。
从图3可以看出,水稻G1期的位置在图中横坐标45.10处,大豆的G1期位置在135.00处,9901柳的G1期位置在74.32处,海柳1号的G1期位置在74.11处。从水稻推算9901柳的基因组大小为440/45.10×74.32=725.1 Mb;推算海柳1号的基因组大小为440/45.10×74.11=723.0 Mb。从大豆推算9901柳的基因组大小为1 150/135×74.32=633.1 Mb;推算海柳1号的基因组大小为1 150/135×74.11=631.3 Mb。
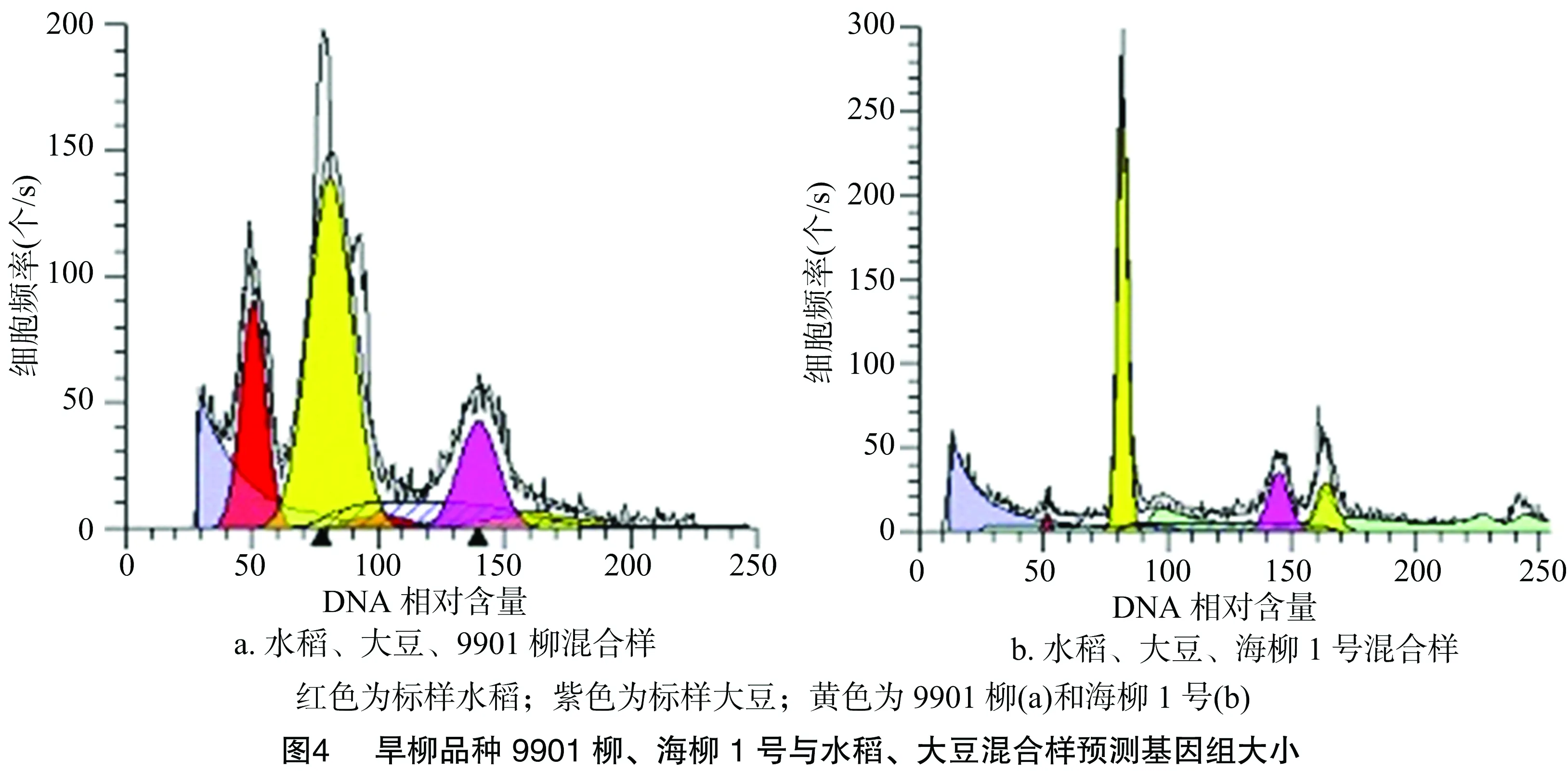
2.2.3 水稻、大豆、9901柳、海柳1号混合样比较 试验同时用流式细胞术对水稻、大豆与9901柳或海柳1号三者的混合样进行测定,并根据测定图计算其基因组大小值。
从图4-a可以看出,水稻G1期位置在图中横坐标 50.56处,大豆G1期位置在139.83处,9901柳的基因组介于两者之间,峰值在81.02处。从水稻推算9901柳的基因组大小为440/50.56×81.02=705.1 Mb,从大豆推算沿江柳的基因组大小为1 150/139.83×81.02=666.3 Mb。
从图4-b可以看出,水稻G1期位置在图中横坐标 51.28处,大豆G1期位置在144.61处,海柳1号的峰值在81.87处。从水稻推算海柳1号的基因组大小为440/51.28×81.87=702.5 Mb, 从大豆推算海柳1号的基因组大小为1 150/144.61×81.87=651.1 Mb。
3 讨论与结论
柳树是我国主要的乡土树种之一,在我国有着悠久的栽培历史。多倍体现象在柳树的物种形成中非常普遍,在已知染色体数目的65种柳树中,2倍体有16种、3倍体有10种、4倍体有43种、5倍体有3种、6倍体有10种、8倍体有7种、10倍体有3种、12倍体有1种[2]。了解柳树的倍性对其重要性状的数量性状座位分析以及全基因组测序有着重要意义,本研究利用流式细胞术以已知二倍体杞柳为对照,测定出旱柳3个品种海柳1号、9901柳和沿江柳均为四倍体。
由于倍性的关系,使得柳树基因组大小差距较大,已知柳树基因组的大小在342~841 Mb之间,如三蕊柳(Salixtriandra,二倍体)的基因组大小为386 Mb[18],簸箕柳(Salixsuchowensis,二倍体)的基因组大小为425 Mb[19],垂柳(Salixbabylonica)的基因组大小为748 Mb[20],白柳(Salixalba,四倍体)的基因组大小为807 Mb[18]。本研究中,笔者运用流式细胞术将旱柳3个品种(沿江柳、9901柳、海柳1号)与基因组大小已知的水稻(基因组大小为440 Mb)和大豆(基因组大小为1 150 Mb)分别进行了对比测定,单液测定结果显示旱柳的基因组大小为605.6~725.1 Mb,混合液测定结果显示的旱柳的基因组大小为651.1~705.1 Mb。综合以上数据,推测旱柳的基因组大小应该在670 Mb左右。
猜你喜欢
江西农业(2021年2期)2021-03-19 04:31:36
农民致富之友(2019年26期)2019-10-11 03:39:10
天津师范大学学报(自然科学版)(2019年4期)2019-09-17 00:59:10
山东林业科技(2018年4期)2018-09-01 08:15:20
作文周刊·小学四年级版(2018年12期)2018-04-10 12:07:56
江苏农业科学(2017年16期)2017-10-27 12:48:11
现代农业科技(2017年8期)2017-06-10 09:32:18
现代农业科技(2016年21期)2017-03-06 00:31:54
河北林业科技(2016年4期)2016-03-12 18:17:14
农产品市场周刊(2015年30期)2015-10-31 08:32:06
