
棉花黄萎病菌拮抗酵母菌筛选及鉴定
2018-12-07 07:15文雨婷孙燕飞雷勇辉
河南农业科学 2018年11期
文雨婷,张 媱,李 杨,孙燕飞,雷勇辉,王 翀
(1.石河子大学 生命科学学院,新疆 石河子 832003; 2.石河子大学 农学院,新疆 石河子 832003;3.新疆出入境检验检疫局,新疆 乌鲁木齐 830063)
棉花是我国重要的经济作物之一,在我国的农业经济中占有举足轻重的地位。新疆是我国最大的棉花产区,随着新疆棉花生产的规模化和棉花轮作模式的普及化[1],棉花黄萎病的发生变得更加迅猛。棉花黄萎病的发病期明显提前,并且危害显著加重,已经严重地制约新疆棉花生产的可持续发展。
棉花黄萎病是由大丽轮枝菌(Verticilliumdahliae)引起的一种土传真菌病害,病菌具有分布广、危害重、寄主范围宽、传播途径多、存活时间久等特点[2-3],现阶段缺乏有效防治棉花黄萎病的手段。除了对棉花黄萎病抗性的研究,生产中多使用化学农药防治棉花黄萎病,但该方法存在严重的环境破坏问题[4-5],生物防治可降低甚至避免这些危害。前人已筛选得到许多对棉花黄萎病菌有较好防治作用的生防菌株,主要有木霉(Trichoderma)[6]、芽孢杆菌(Bacillus)[7]、假单胞菌(Pseudomonas)[8]、链霉菌(Streptomyces)[9]、毛壳菌(Chaetomium)[10]等。一些研究证明,从植物内部、表皮及根际分离出的微生物可以在一定程度上直接或间接降低植物病原菌的危害[11-13],而酵母菌是植物附生微生物的重要组成部分,较其他微生物而言具有遗传稳定、抑菌谱广、抗逆性强、安全性高等特点,因此常被用作病原菌的生物控制剂[14-16]。目前,利用酵母菌防治棉花黄萎病的研究鲜有报道,同时由于生防菌剂具有明显的地域性,跨地域使用往往效果欠佳。新疆地区含有丰富的酵母菌资源,因此,以新疆地区分离的酵母菌株为对象,筛选能够抑制棉花黄萎病菌生长的酵母菌,为利用酵母菌防治棉花黄萎病提供理论依据和实践基础。
1 材料和方法
1.1 材料
菌株:石河子大学生命科学学院微生物生态实验室保存的黄萎病菌VD911及从新疆地区果园、甜菜地、活性污泥等土壤中或水果表皮分离纯化获得的230株酵母菌。
培养基:马铃薯葡萄糖琼脂(PDA)培养基、酵母浸出粉胨葡萄糖(YEPD)培养基、糖类发酵培养基、碳源同化培养基。
1.2 拮抗酵母菌的筛选
采用平板对峙法[17-18],在PDA固体培养基中心放置直径为8 mm的黄萎病菌菌饼,将待筛酵母菌点接于距菌饼2.5 cm处,以不接种酵母菌的培养基为空白对照,观察酵母菌附近黄萎病菌菌丝生长的情况。琼脂块法参照文献[19],挑取直径为7 mm、有抑菌活性的酵母菌菌饼,分别接种于孢子浓度为104、105、106cfu/mL(挑取黄萎病菌菌丝于PDA液体培养基中,培养7 d,用4层纱布过滤,收集孢子滤液,计数待用)的黄萎病菌平板上,7 d后观察拮抗酵母菌的抑菌效果并测量其抑菌圈直径,每组试验重复3次。
1.3 拮抗酵母菌形态观察及生理生化特性检测
将筛选获得的酵母菌株在 YEPD 培养基平板上划线接种,28 ℃培养 2~5 d后进行菌落形态观察,记录菌落特征;将菌种接种于YEPD 液体培养基中,28 ℃振荡培养 24 h ,取样在显微镜下观察菌体形态。酵母菌的生理生化鉴定方法参照《The yeasts: A taxonomic study》[20]。
1.4 拮抗酵母菌系统发育分析
将筛选获得的酵母菌接种于20 mL YEPD 液体培养基中,28 ℃培养过夜,离心收集菌体,用十六烷基三甲基溴化铵(Cetyl trimethyl ammonium bromide,CTAB)法提取DNA后进行26S rDNA 的PCR扩增。扩增采用酵母26S rDNA通用引物NL-1:5′-GCATATCAATAAGCGGAGGAAAAG-3′和NL-4:5′-GGTCCGTGTTTCAAGACGG-3′。PCR体系及程序参照文献[21]并加以改进。扩增体系共25 μL:10×TaqBuffer 2.5 μL,10 mmol/L dNTP 2 μL,25 mmol/L MgCl21.5 μL,NL-1 1 μL,NL-4 1 μL,DNA 2 μL,TaqDNA聚合酶0.625 μL,ddH2O 14.375 μL。PCR 程序为:94 ℃ 10 min;94 ℃ 1 min,56 ℃ 45 s,72 ℃ 1 min,30个循环;72 ℃ 10 min。将扩增出的26S rDNA序列送上海生工生物工程有限公司测序。登录NCBI网站(http://www.ncbi.nlm.nih.gov/)用BLAST程序进行序列比对,下载序列,通过MEGA 6.0软件进行分析并构建进化树。
2 结果与分析
2.1 拮抗酵母菌筛选
通过对峙培养,从230株酵母菌筛选获得ZHB39、19-10、F3、S4B2-13、T1、MLY1共6株效果较好的拮抗酵母菌株(图1)。随后在不同孢子浓度的黄萎病菌平板上接种拮抗酵母菌,对比其抑菌圈大小发现,随着黄萎病菌孢子浓度的降低,拮抗酵母菌的抑菌活性增加。当黄萎病菌孢子浓度为104cfu/mL时,拮抗酵母菌的抑菌圈直径最大,此时6株拮抗酵母菌的抑菌圈直径分别为12.50、13.83、16.33、25.17、21.83、11.17 mm。其中,S4B2-13的抑菌活性显著优于其他5株酵母菌,T1的抑菌活性显著优于MLY1、ZHB39、19-10和F3,F3的抑菌活性显著优于MLY1、ZHB39和19-10(表1)。但由图1可知,S4B2-13和T1产生的抑菌圈内仍有少许黄萎病菌菌丝生长,而F3形成的抑菌圈中无任何黄萎病菌菌丝的生长。
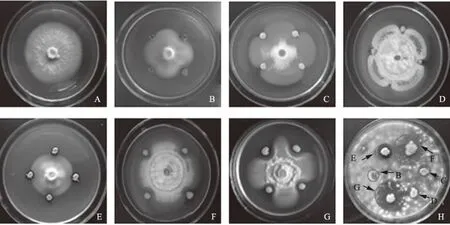
A:空白对照;B:接种拮抗酵母菌株MLY1;C:接种拮抗酵母菌株ZHB39;D:接种拮抗酵母菌株19-10;E:接种拮抗酵母菌株F3;F:接种拮抗酵母菌株T1;G:接种拮抗酵母菌株S4B2-13;H:6株拮抗酵母菌产生的抑菌圈大小图1 拮抗酵母菌筛选
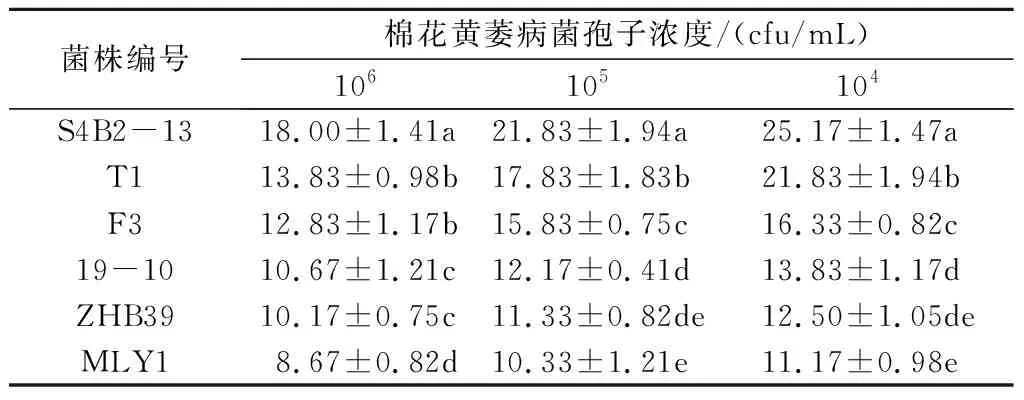
表1 拮抗酵母菌的抑菌圈直径 mm
注:同列不同字母代表不同菌株的抑菌圈直径差异显著(P<0.05)。
2.2 拮抗酵母菌的形态特征
本试验共筛选获得6株拮抗棉花黄萎病菌的酵母菌,其形态特征各不相同(图2、表2)。其中ZHB39、T1和MLY1菌落边缘整齐,而19-10边缘不整齐,F3边缘生出类似根状的分支,S4B2-13呈丝状;ZHB39和T1菌落隆起呈枕状,19-10呈脐凹状,F3隆起呈脐凸状,S4B2-13隆起呈扁平状,MLY1稍拱起;ZHB39、19-10、F3、S4B2-13和T1菌落不透明,而MLY1呈胶状。ZHB39和F3的菌体形态呈椭圆形,19-10呈圆形,S4B2-13呈鞋形,T1呈柠檬形,MLY1呈长杆状。因此,从菌落和菌体的差异上认为筛选出的6个拮抗菌株属于不同的酵母菌。
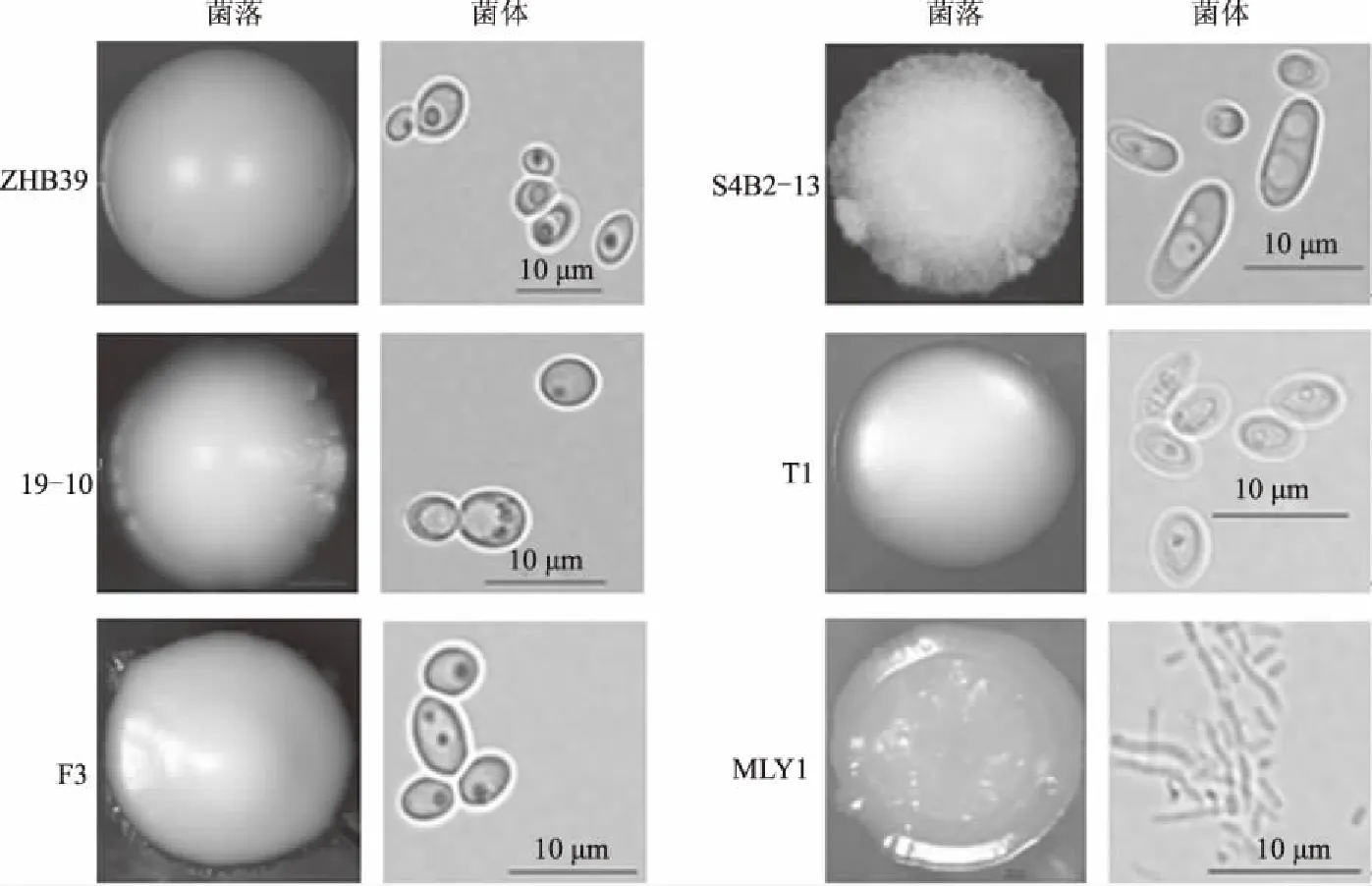
图2 拮抗酵母菌的菌落和菌体形态

表2 拮抗酵母菌菌落菌体形态及相近物种序列相似性分析
2.3 拮抗酵母菌系统发育分析
对6株拮抗酵母菌进行26S rDNA 的PCR扩增,获得大小为 500~600 bp的目的片段,登录NCBI网站用BLAST程序进行序列比对,发现ZHB39、19-10、F3、S4B2-13、T1和MLY1的相近物种分别为美极梅奇酵母(Metschnikowiapulcherrima)、Kazachstaniazonata、美极梅奇酵母、威克汉姆酵母(Wickerhamomycesanomalus)、葡萄汁有孢汉逊酵母(Hanseniasporauvarum)和Barnettozymacalifornica,其遗传相似性均大于98%,且E值均为0(表2)。对拮抗酵母菌和其相似物种利用MEGA 6.0软件构建系统发育树,如图3所示,19-10与Kazachstania属的酵母菌聚集于一个大的遗传分支,但与该属中的标准株距离较远;F3和ZHB39与梅奇酵母属(Metschnikowia)的酵母菌聚集于一个大的遗传分支,并且F3和ZHB39又形成一个相对独立的遗传分支,亲缘关系较近,但与该属的标准株距离较远;T1与汉逊酵母属(Hanseniaspora)的酵母菌聚集于一个大的遗传分支,并与葡萄汁有孢汉逊酵母在该遗传分支中进一步形成一个相对独立的遗传分支;MLY1与Barnettozyma属的酵母菌聚集于一个大的遗传分支,但与该属中的标准株距离较远;S4B2-13与威克汉姆酵母聚集于一个遗传分支。因此,初步认为ZHB39、19-10、F3、S4B2-13、T1和MLY1这6株拮抗酵母菌分别为梅奇酵母菌(Metschnikowiasp.)、Kazachstaniasp.、梅奇酵母菌、威克汉姆酵母、葡萄汁有孢汉逊酵母和Barnettozymasp.。
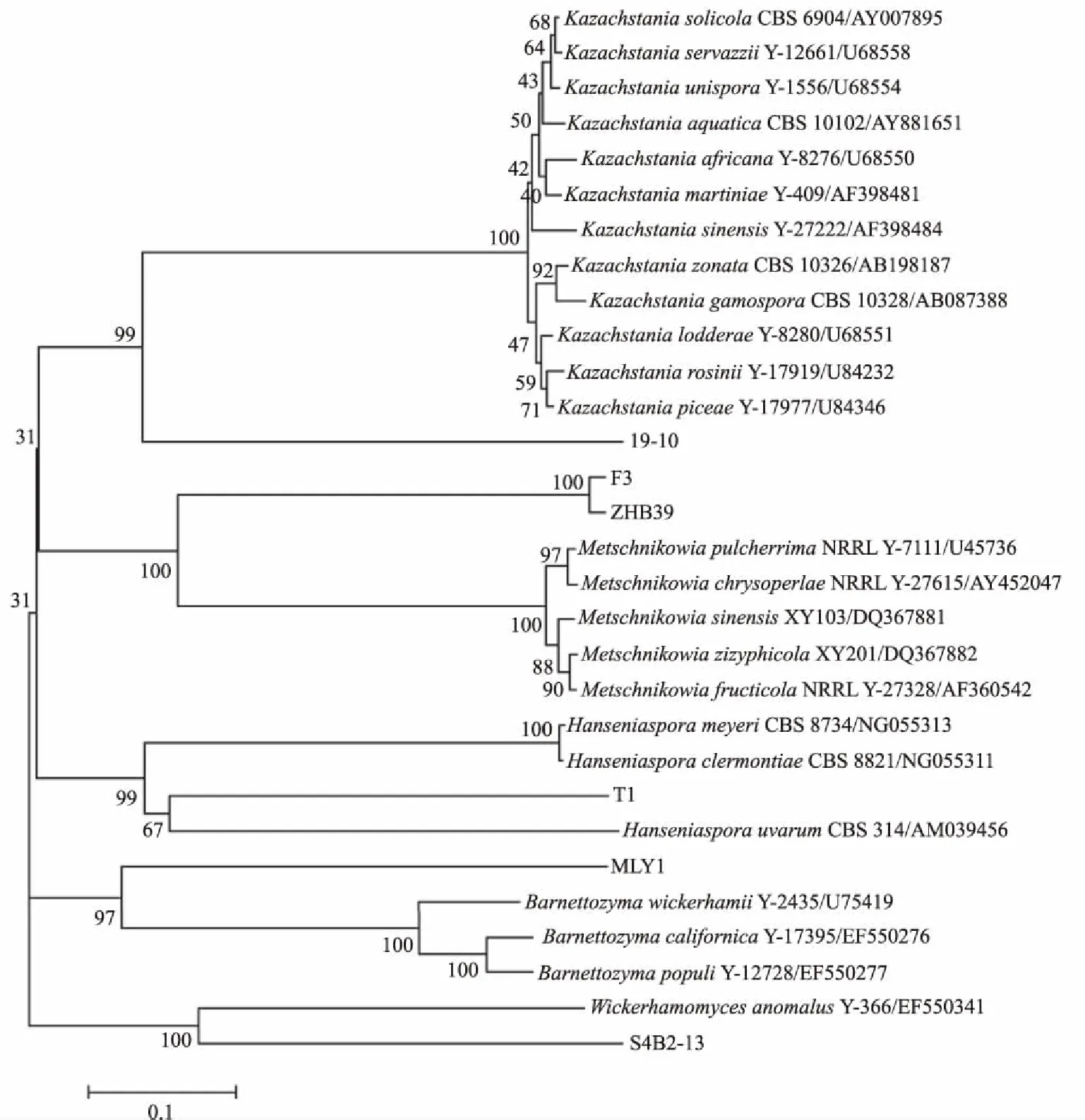
图3 基于26S rDNA序列构建的拮抗酵母菌株的系统发育树
2.4 拮抗酵母菌的生理生化特性
根据6株酵母菌的系统发育分析结果,并参照《The yeasts: A taxonomic study》[20]手册,进一步对6株酵母菌进行相关生理生化特性检测,对其种属地位进行最终确定。糖发酵试验表明,6株拮抗酵母菌均可利用葡萄糖进行发酵,但S4B2-13还能利用蔗糖进行发酵,19-10还能利用蔗糖、麦芽糖和棉籽糖进行发酵(表3)。碳源利用试验结果表明(表4),ZHB39和F3均能利用葡萄糖、蔗糖、半乳糖、海藻糖、乙醇、甘油、D-甘露醇、麦芽糖、松三糖、甲基-吡喃葡萄糖、纤维二糖、水杨苷、D-山梨醇、琥珀酸钠、柠檬酸盐、葡萄糖酸盐、D-葡萄糖胺和鲸蜡烷等碳源进行同化作用,19-10能利用葡萄糖、菊粉、蔗糖、棉籽糖和麦芽糖等碳源进行同化作用,S4B2-13能利用葡萄糖、蔗糖、棉籽糖、海藻糖、乙醇、甘油、赤藓糖醇、麦芽糖、松三糖、甲基-吡喃葡萄糖、可溶性淀粉、纤维二糖、水杨苷、D-甘露醇、D-山梨醇、乳酸钠、琥珀酸钠和柠檬酸盐等碳源进行同化作用,T1能利用葡萄糖、纤维二糖和水杨苷等碳源进行同化作用,MLY1能利用葡萄糖、乙醇、甘油、D-甘露醇、纤维二糖、水杨苷、L-山梨糖、D-木糖、乳酸钠、琥珀酸钠和葡萄糖酸盐等碳源进行同化作用。温度、高渗、尿素酶、无维生素等生理生化特性检测结果表明(表5),除MLY1最适生长温度为28 ℃外,其余酵母菌均为38 ℃;6株酵母菌均能在50%葡萄糖培养基中生长,不能在无维生素培养基中生长;ZHB39、19-10、F3能产生弱的尿素酶反应,MLY1产生强尿素酶反应,S4B2-13和T1不能进行尿素酶反应。经鉴定,19-10是Kazachstaniasp.,S4B2-13是威克汉姆酵母,T1是葡萄汁有孢汉逊酵母,MLY1是Barnettozymacalifornica,ZHB39和F3是美极梅奇酵母。6株拮抗酵母菌的一些生理生化特性如温度、尿素酶反应等与《The yeasts: A taxonomic study》[20]上记录的略有差异,推测可能是同一个种不同亚种之间的区别。
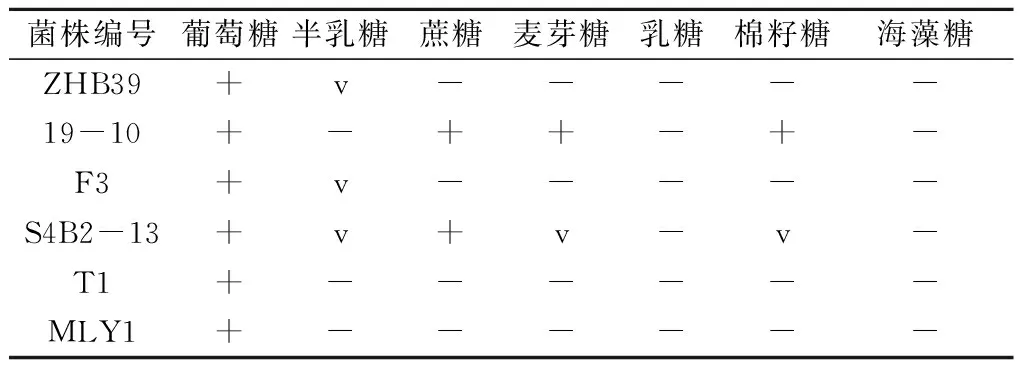
表3 拮抗酵母菌糖发酵检测结果
注:+代表发酵;v代表部分发酵,部分不发酵;-代表不发酵。
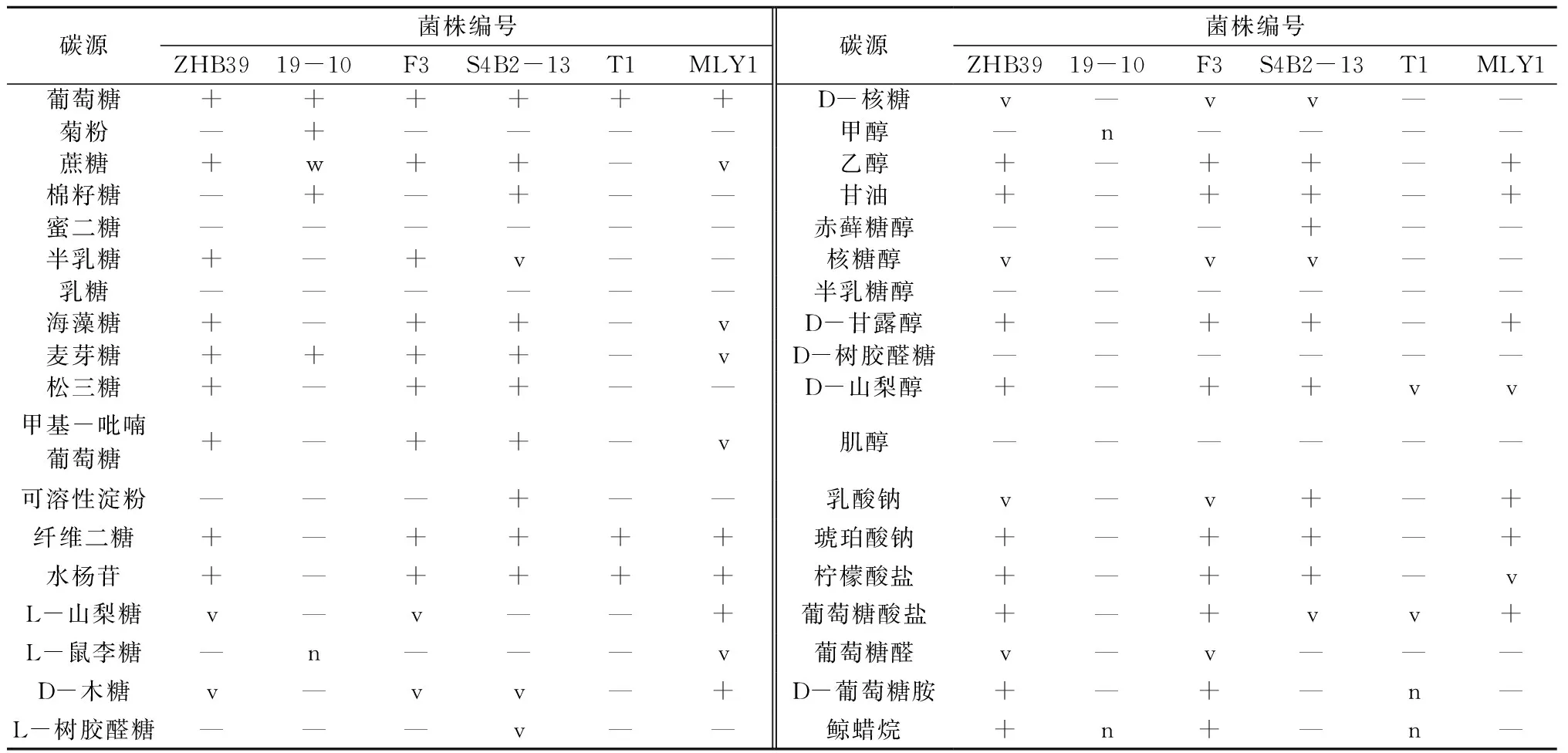
表4 拮抗酵母菌碳源利用试验结果
注:+代表强同化;w代表弱同化;v代表部分同化,部分不同化;—代表不同化;n代表无数据。

表5 拮抗酵母菌部分生理生化特性检测结果
注:+代表生长或反应;w代表弱生长或弱反应;-代表不生长或不反应。
3 结论与讨论
本研究共获得6株拮抗棉花黄萎病菌的酵母菌,分别为ZHB39、19-10、F3、S4B2-13、T1和MLY1,经鉴定,19-10是Kazachstaniasp.,S4B2-13是威克汉姆酵母(Wickerhamomycesanomalus),T1是葡萄汁有孢汉逊酵母(Hanseniasporauvarum),MLY1是Barnettozymacalifornica,ZHB39和F3是美极梅奇酵母(Metscgnikowiapulcherrima)。6株拮抗酵母菌在黄萎病菌孢子浓度为104cfu/mL时,抑菌圈直径最大,分别为12.50、13.83、16.33、25.17、21.83、11.17 mm,均具有较好的抑菌能力。
近年来,有许多关于酵母菌防治植物病害的研究报道,尤其在酵母菌对水果采后病害的防治中,靳莎莎等[22]筛选出1株抑制草莓灰霉病和根霉病的拮抗酵母菌卡利比克毕赤酵母菌(Pichiacaribbica),并且发现在一定条件下,灰霉病和根霉病的腐烂率可被完全抑制;高云慨等[23]研究表明,季也蒙毕赤酵母菌(Meyerozymaguilliermondii)可有效抑制芒果炭疽病。已报道的拮抗酵母菌主要有红酵母菌(Rhodotorulasp.)[24]、葡萄汁有孢汉逊酵母菌[25]、梅奇酵母菌(Metscgnikowiasp.)[26]、柠檬克勒克酵母菌(Kloeckeraapiculata)[27]、隐球酵母菌(Cryptococcussp.)[28]、假丝酵母菌(Candidasp.)[29]等。而关于Kazachstaniasp.和Barnettozymacalifornica防治植物病害的研究几乎没有报道,威克汉姆酵母菌多用于发酵,也几乎没有关于防治植物病害的研究,本研究发现Kazachstaniasp.、威克汉姆酵母菌、Barnettozymacalifornica对于棉花黄萎病菌有一定的抑制作用,这对酵母菌防治植物病害的研究具有重要意义。在酵母抑制植物病原菌的研究中,张建等[30]发现,德巴利酵母菌(Debaryomycessp.)I8对蟠桃根腐病菌的抑菌圈直径为17.20 mm;任少东等[31]发现,拮抗酵母菌对致病菌的平均抑菌圈直径为20~30 mm。在生防菌抑制棉花黄萎病菌的研究中,陈丽华等[32]试验结果表明,内生芽孢杆菌LH-L3对棉花黄萎病菌的抑菌圈直径为25.00 mm;刘海洋等[33]试验结果表明,甲基营养型芽孢杆菌AL7的脂肽粗提取物对棉花黄萎病菌的抑菌圈直径为36.1 mm;金利容等[34]试验结果表明,内生恶臭假单胞菌HB3S-20菌株对棉花黄萎病菌的抑菌圈直径为13.70 mm。本试验筛选出的拮抗酵母菌S4B2-13对棉花黄萎病菌的抑菌圈直径为25.17 mm,T1对棉花黄萎病菌的抑菌圈直径为21.83 mm,均具有较强的抑菌效果,可进一步进行盆栽试验验证。
本研究共获得6株拮抗棉花黄萎病菌的酵母菌株,其中ZHB39和F3均属于美极梅奇酵母,但两者间的抑菌能力有差异,并且菌落形态也不相同,推测ZHB39和F3可能属于同种间不同亚种,因此,下一步应对其抑制黄萎病菌的有效成分进行检测和确定。19-10的生理生化特性与Kazachstaniasp.中相似种的生理生化特性均有差异,故很有可能是新菌种,还需进一步进行5.8S-ITS序列分析和更多理化因子测定。对于拮抗酵母的拮抗因子和抑菌效果,还需要进一步进行代谢产物分离检测和相应的盆栽及大田试验。
猜你喜欢
作物学报(2022年9期)2022-07-18
环境工程技术学报(2022年3期)2022-06-05
昆明医科大学学报(2022年1期)2022-02-28
中国妇幼健康研究(2022年1期)2022-02-25
农业科技与信息(2021年4期)2021-12-05
中国土壤与肥料(2021年5期)2021-12-02
昆钢科技(2021年6期)2021-03-09
中国棉花(2021年11期)2021-03-05
军事文摘·科学少年(2021年1期)2021-02-04
天津科技(2020年4期)2020-05-09
