
桉树焦枯病菌SIT转运蛋白的鉴定与表达
2018-12-06 05:24刘宏毅陈慧洁李慧敏叶小真冯丽贞郭朦朦
西北农林科技大学学报(自然科学版) 2018年11期
刘宏毅,陈慧洁,李慧敏,叶小真,冯丽贞,郭朦朦
(福建农林大学 a 林学院,b 金山学院,福建 福州 350002)
在大多数原核生物和所有真核生物中,铁元素是各种生命活动的必需元素。地壳中铁元素储量丰富,但其难溶性大大限制了真菌对铁的直接吸收。在生境中铁元素含量较低时,大多数微生物和部分植物通过分泌铁载体与Fe3+结合为铁载体-铁化合物来获取铁元素[1]。大多数真菌存在4种异羟肟酸型铁载体,分别为镰孢氨酸、粪生素、铁色素和罗丹明酸,其主要由非核糖体肽合成酶(NRPSs,non-ribosomal peptide synthetases)来合成[2]。铁载体-铁转运蛋白(Siderophore iron transporter,SIT)属于MFS(Major facilitator superfamily)超家族,在缺铁生境下通过转运铁载体-铁化合物为微生物的生长发育提供铁元素。SIT转运蛋白具有典型的MFS结构域,家族成员大多数由400~600个氨基酸残基组成,N端和C端均位于胞内,MFS蛋白的二级结构大多含有12个跨膜α螺旋,仅少数含有6,14或24个[3-4],目前已知的SIT转运蛋白均含有14个跨膜α螺旋,多出的2个跨膜螺旋是由胞内中间的环状区(loop)插入膜中产生的。因此,虽然少数蛋白成员含有的跨膜α螺旋数目不同,但是并不影响MFS超家族整体的蛋白折叠方式,即MFS折叠[5]。MFS转运蛋白的转运机制由摇杆开关和门控运输理论组成[6-8],MFS蛋白的N端或C端的跨膜螺旋围绕着底物结合位点一侧关闭一侧开放以进行转运。SIT转运蛋白作为铁载体-铁化合物的运输载体发挥着重要作用,如禾谷镰刀菌(Fusariumgraminearum)FgSit1基因编码的SIT转运蛋白负责调控铁载体-铁化合物FC-Fe3+的吸收[9],构巢曲霉(Aspergillusfumigatus)的SIT转运蛋白MirB与铁载体-铁化合物三乙酰镰孢氨酸的转运有关[10],酿酒酵母(Saccharomycescerevisiae)Arn1p转运蛋白通过转运生境中的铁载体-铁化合物来获取铁元素[11]。
桉树焦枯病(Calonectriapseudoreteaudii)是热带和亚热带地区桉树种植区危害最为严重的病害之一,严重威胁桉树产业的发展[12]。桉树焦枯病由丽赤壳属(Calonectria)真菌引起,其无性态为帚梗柱孢属(Cylindrocladium)真菌[13]。据统计,Calonectria现有集群13个共71种,其中Calonectriapseudoreteaudii是福建省内发现最早、分布最广、致病力最强的病原菌株[14-16]。有研究发现,桉树焦枯病菌SIT转运蛋白在焦枯病菌侵染桉树48 h时上调表达,可能在病原菌侵染寄主的过程中,CpSit1基因通过调控铁载体-铁化合物的转运完成铁元素的摄入,进而协助焦枯病菌在桉树中的定植[17]。为了解CpSit1基因在桉树焦枯病菌中是否为单拷贝基因,本研究通过多种生物信息学软件在全基因组范围内对焦枯病菌SIT蛋白家族进行鉴定,并对其编码基因、结构域、跨膜螺旋、亚细胞定位等进行分析和功能预测,进一步通过构建系统发育树进行多菌种比较分析,并通过qPCR分析SIT转运蛋白在焦枯病菌侵染桉树48 h后的表达情况,以期为揭示SIT转运蛋白家族在焦枯病菌铁元素摄入机制及铁载体-铁转运蛋白在焦枯病菌侵染桉树过程中所扮演的角色奠定基础。
1 材料与方法
1.1 基因组序列来源
桉树焦枯病菌(Calonectriapseudoreteaudii,菌株:YA51)基因组NCBI检索号为MOCD01000000。禾谷镰刀菌(Fusariumgraminearum,菌株:PH-1)、粗糙脉胞菌(Neurosporacrassa,菌株:OR74A)、稻瘟菌(Magnaportheoryzae,菌株:70-15)、灰葡萄孢(Botrytiscinerea,菌株:B05-10)的基因组数据及蛋白序列数据,均下载自Broad Institute(www.broadinstitute.org/)数据库。
1.2 试验方法
1.2.1 SIT转运蛋白的鉴定 从NCBI数据库中下载所有真菌SIT转运蛋白的蛋白序列,通过MAFFT软件进行多重比较;然后通过HMMER软件(http://hmmer.org/)的hmmbuild功能构建隐马可夫(HMM,Hidden Markov Model)模型,用hmmsearch功能将其与桉树焦枯病菌、禾谷镰刀菌、粗糙脉胞菌、稻瘟菌和灰葡萄孢的蛋白序列库进行比对,E值设定为1×10-10;进一步用hmmsearch分析结果再次构建隐马可夫模型后用hmmsearch进行迭代搜索;最后通过与TCDB(Transporter Classification Database, http://www.tcdb.org/)数据库进行Blast比对,最终获得这5个菌种的SIT转运蛋白。
1.2.2 结构域预测 采用InterPro(http://www.ebi.ac.uk/interpro/)在线分析功能进行SIT转运蛋白的结构域预测。
1.2.3 跨膜螺旋预测 采用TMHMM Server v.2.0(http://www.cbs.dtu.dk/services/TMHMM/)对SIT转运蛋白的跨膜螺旋进行预测。
1.2.4 亚细胞定位预测 采用WoLF PSORT在线工具(https://wolfpsort.hgc.jp/)对SIT转运蛋白进行亚细胞结构定位预测。
1.2.5 系统发育分析 采用mafft软件[18]对禾谷镰刀菌、稻瘟菌、灰葡萄孢、粗糙脉胞菌和桉树焦枯病菌的SIT转运蛋白进行多重比对,以禾谷镰刀菌的DHA14(drug:H+antiporter 14 spanner)蛋白TRI102为外群,然后用Gblock软件去除冗余序列,最后用Neighbor-Joining法通过MEGA7.0[19]构建其系统发育树。bootstrap检测值设为1 000次,其余参数均为系统默认值,对重复率小于50%的分支进行合并。
1.2.6 SIT基因表达分析 基于焦枯病菌侵染桉树48 h的数字基因表达谱数据(Digital Gene Expression profiling,DGE),获得桉树焦枯病菌SIT基因的转录表达情况[20],并将其表达情况标注在系统发育树中。采用与DGE分析相同的方法制备样品,然后进行总RNA提取、PCR反转录及实时荧光PCR,其中总RNA提取、反转录PCR(Reverse transcription PCR,RT-PCR)、实时荧光定量PCR(Quantitative real-time PCR,qRT-PCR)试剂盒均购自天根生化科技(北京)有限公司。采用Talent qPCR PreMix试剂盒进行qRT-PCR,反应体系为:2×Talent qPCR Mix 10.0 μL,上游引物0.6 μL,下游引物0.6 μL,cDNA模板1.0 μL,50×ROX Reference Dye 0.4 μL,并加RNase-Free ddH2O至20 μL。采用Beacon Design 7.9软件进行引物设计(表1),委托生工生物工程有限公司合成。采用2-ΔΔCt法计算焦枯病菌SIT基因的表达量。

表1 桉树焦枯病菌SIT转运蛋白的引物Table 1 Primers of SIT transporters in Calonectria pseudoreteaudii
2 结果与分析
2.1 SIT转运蛋白的鉴定
通过对桉树焦枯病菌全基因组进行hmmsearch分析,从14 355条序列中共鉴定出16个SIT转运蛋白(表2),约占总编码蛋白的0.11%。序列分析发现,其蛋白序列长度大多数为400~603个氨基酸,编码这些蛋白序列的ORF(Open reading frame)阅读框位于不同的基因组支架(Scaffold)上。桉树焦枯病菌SIT转运蛋白的蛋白序列与TCDB数据库比对结果(表2)显示,其主要分为13个铁-铁载体转运蛋白(Ferri-siderophore transporter,TCID:2.A.1.16.7)、2个铁载体-铁:H+同向转运蛋白(Siderophore-iron:H+symporter,TCID:2.A.1.16.1)和1个铁载体-铁转运蛋白(Siderophore iron transporter,TCID:2.A.1.16.6)。亚细胞定位预测结果(表2)显示,除CpSit13转运蛋白在内质网上外,其余均在细胞膜上。
2.2 结构域及跨膜螺旋分析
保守结构域分析发现,16个桉树焦枯病菌SIT转运蛋白均为MFS结构域(图1),其InterPro ID为IPR020846。SIT蛋白结构域长短不一,长度为24~524个氨基酸,其中CpSit4、CpSit8、CpSit10、CpSit11和CpSit13分为2段;CpSit1和CpSit5分为3段。CpSit1的2段长度较为接近的结构域的一致性为21%,CpSit5 的2段长度较为接近的结构域的一致性为23%,说明CpSit1和CpSit5转运蛋白的结构域在进化晚期通过单个结构域复制而来的可能性较低。

表2 桉树焦枯病菌SIT蛋白的相关信息Table 2 Information of siderophore-iron transporters in Calonectria pseudoreteaudii
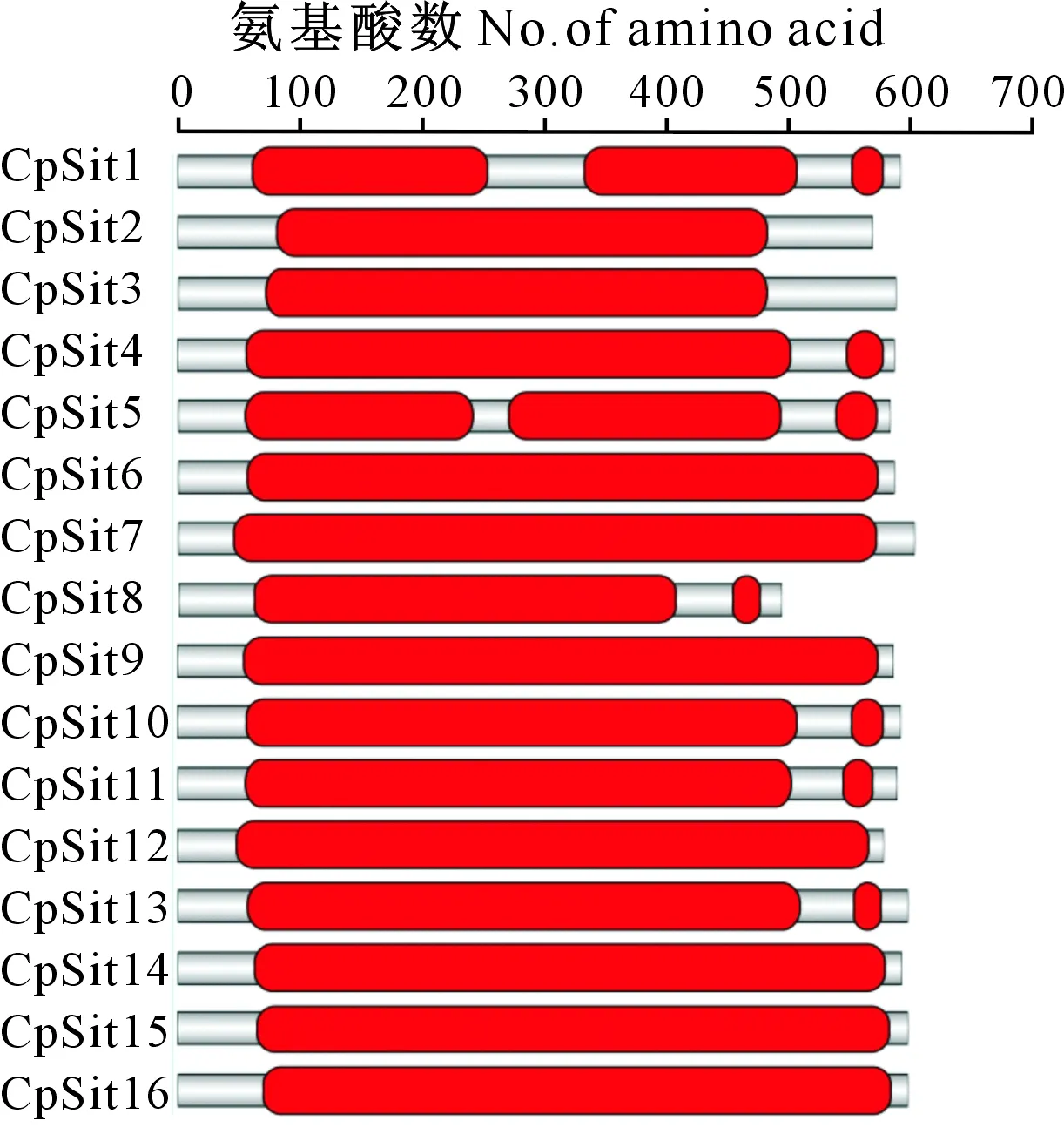
图1 桉树焦枯病菌SIT转运蛋白的结构域Fig.1 Conserved domains of Siderophore-iron transporters in Calonectria pseudoreteaudii
TMHMM分析(表2)发现,CpSit3含有12个跨膜螺旋,CpSit5、CpSit11和CpSit15均含有13个,CpSit7含有11个,CpSit8含有10个,其余均含有14个跨膜螺旋。此外,CpSit8的蛋白长度及结构域长度明显小于其他SIT转运蛋白,其跨膜螺旋数仅为10个,与典型SIT蛋白的跨膜螺旋数量不符,这可能是基因复制过程中结构遗失或基因组拼装错误所致。虽然部分桉树焦枯病菌SIT转运蛋白的跨膜α螺旋数目不尽相同,但并不影响整体的蛋白折叠方式,其跨膜螺旋较为均匀地分布在N端和C端,为典型的MFS折叠。因此推测SIT蛋白与底物结合后,其N端或C端的跨膜螺旋围绕着底物结合位点一侧关闭一侧开放进行转运。以上结果表明,桉树焦枯病菌SIT转运蛋白结构特征较为保守,可能具有活性和功能。
2.3 不同物种间SIT转运蛋白的比较
通过hmmsearch共从禾谷镰刀菌、稻瘟菌、粗糙脉胞菌、灰葡萄孢和桉树焦枯病菌中发现34个SIT转运蛋白,其中禾谷镰刀菌8个,稻瘟菌3个,粗糙脉胞菌2个,灰葡萄孢5个,而桉树焦枯病菌有16个SIT转运蛋白,存在明显扩张。对上述SIT蛋白序列进行结构域分析,发现其结构域均为MFS结构域。
通过构建系统发育树(图2),发现其分支聚为3大类,分别为铁载体-铁:H+同向转运蛋白、铁载体-铁转运蛋白和外群TRI102蛋白,表明这些SIT转运蛋白具有明显的直系同源关系,来源于共同的祖先基因且在进化中十分保守。桉树焦枯病菌SIT转运蛋白在进化树中分别聚在不同分支中,不存在单拷贝状况。在禾谷镰刀菌、稻瘟菌、粗糙脉胞菌、灰葡萄孢和桉树焦枯病菌中,仅禾谷镰刀菌和焦枯病菌分别含有2个铁载体:H+同向转运蛋白,这种在特定菌种中特有的SIT,可能暗示着与这2个菌种相应的生物学功能存在相关性,但具体情况还有待后续研究。

(1)红色分支为铁载体-铁:H+同向转运蛋白,蓝色分支为铁载体-铁转运蛋白,绿色分支为外群TRI102蛋白;(2)五边形标记部分为桉树焦枯病菌SIT蛋白转录表达情况,黄色代表上调,褐色代表下调,灰色代表表达不显著(1)The red branch represents the Siderophore-iron:H+ Symporter,the blue branch represents the Siderohore iron transporter and the green color represents the outgroup TRI102 protein;(2)The pentagon is the expression of SIT protein in Calonectria pseudoreteaudii,the yellow represents up-regulation,the brown represents down-regulation and the gray represents false图2 5种真菌SIT转运蛋白的系统发育树Fig.2 Phylogenic analysis of Siderophore-iron transporters in 5 species
2.4 SIT基因表达分析
由表3可知,焦枯病菌侵染桉树48 h后,CpSit1、CpSit3、CpSit5、CpSit8、CpSit9、CpSit10、CpSit11、CpSit14、CpSit15和CpSit16 10个基因上调表达。

表3 桉树焦枯病菌SIT基因的相对转录表达量Table 3 Relative transcription levels of SIT genes in Calonectria pseudoreteaudii
结合系统发育树聚类分析结果(图2)可知,CpSit11和CpSit13聚在同一分支上,但是CpSit13表达不显著,CpSit11上调且转录量较低;CpSit1和CpSit10,CpSit3、CpSit14和CpSit16,CpSit8和CpSit9,CpSit5和CpSit15分别聚在同一分支上,均为上调表达,且相对分支中其他基因具有较高的转录量。推测CpSit1、CpSit5、CpSit8、CpSit14和CpSit16 5个基因在焦枯病菌侵染桉树的过程中具有较为重要的作用,CpSit3、CpSit9、CpSit10、CpSit11和CpSit15基因的作用可能相对较弱。CpSit2和CpSit4 2个基因下调表达,推测其可能在焦枯病菌侵染桉树的过程中发挥负调控作用。CpSit6、CpSit7、CpSit12和CpSit13 4个基因表达不显著,说明这4个基因在焦枯病菌侵染桉树过程中作用较弱,或在所研究的范围之外发挥其他作用。
为了验证上述基因转录数据的准确性,利用实时定量PCR对其中10个SIT基因进行分析,结果如图3所示。图3显示qPCR结果与转录组的一致性较高,CpSit1、CpSit5、CpSit8、CpSit14和CpSit16均具有较高的表达量,且CpSit2和CpSit4均下调表达。转录分析表明,CpSit1、CpSit5、CpSit8、CpSit14和CpSit16基因可能在焦枯病菌侵染桉树48 h时发挥着较为重要的作用。

图3 10个SIT基因相对转录表达量的比较Fig.3 Comparison of the relative expressions of SIT genes in Calonectria pseudoreteaudii
3 讨论与结论
随着病原真菌基因组测序工作的深入开展,越来越多的SIT转运蛋白获得鉴定,但是其在林木病原真菌中的鉴定尚无相关报道。为弄清SIT转运蛋白在焦枯病菌侵染桉树过程中所起的作用,本研究对桉树焦枯病菌SIT转运蛋白进行鉴定和比较,并对其结构域、跨膜螺旋、进化关系和基因表达等进行了分析,结果表明桉树焦枯病菌共含有16个SIT转运蛋白,分别是13个铁-铁载体转运蛋白、2个铁载体-铁:H+同向转运蛋白和1个铁载体-铁转运蛋白。桉树焦枯病菌SIT转运蛋白的结构域、跨膜螺旋数与典型SIT蛋白的一致性较高,其跨膜螺旋折叠方式为典型的MFS折叠。由于SIT转运蛋白结构的保守性,其可能具有相类似的功能和活性。
作为微生物铁元素的摄入途径之一,铁载体-铁化合物转运途径对微生物的生长发育起着重要作用。如玉米大斑病菌(Setosphaeriaturcica)铁载体合成基因NPS6的突变体在铁饥饿生境下生长几乎受到完全抑制,说明玉米大斑病菌的铁载体-铁化合物的吸收与其生长发育密切相关[21]。除此之外,铁载体-铁化合物的转运还与病原真菌的致病性有关。Hof等[22]研究表明,稻瘟菌铁载体合成酶基因SSM1被敲除后,其致病性明显减弱;Oide等[23]研究表明,玉米小斑病(Cochliobolusheterostrophus)的铁载体合成基因NPS6获得敲除后致病力减弱。但SIT转运蛋白作为铁载体-铁化合物的运输途径,与其致病性是否存在关联尚未明确。有研究表明,轮枝镰孢菌(Fusariumverticillioides)FIR1转运蛋白在低铁条件下上调表达,但与其致病性不存在关联[24]。然而,苹果腐烂病菌(Valsamali)共含有7个SIT转运蛋白,其中3个在苹果腐烂病菌侵染苹果树的过程中发生上调[25]。为了解桉树焦枯病菌是否与其致病性存在关联,本研究基于焦枯病菌侵染桉树48 h转录组的基因表达谱,对16个SIT基因的表达情况进行分析,发现其中10个基因上调表达、2个下调表达、4个表达不显著。通过系统发育树聚类分析发现,虽然部分焦枯病菌SIT基因聚集在同一分支上,但其表达量存在较为明显的差异。其中,CpSit1、CpSit5、CpSit8、CpSit14和CpSit16相对分支中的其他基因具有较高的转录量,由此推测CpSit1、CpSit5、CpSit8、CpSit14和CpSit16等5个基因在焦枯病菌侵染桉树的过程中发挥着较为重要的作用,其余基因的作用可能相对较弱,或在所研究的范围之外发挥其他作用。为验证基因表达谱的准确性,本研究通过qPCR对其中10个SIT基因进行验证,与基因表达谱具有较高的一致性,其中CpSit1、CpSit5、CpSit8、CpSit14和CpSit16等5个基因均有较高的表达量。
综上所述,SIT转运蛋白很可能与桉树焦枯病菌基因的致病性密切相关。由于铁元素摄取的重要性,后续可将SIT转运蛋白作为毒力因子,通过基因敲除等分子生物学手段对其功能进行验证,了解其在相应生物学过程中的具体作用,为抗焦枯病菌药物的研制提供靶标基因。
猜你喜欢
中华实验眼科杂志(2022年8期)2022-11-15
当代水产(2022年1期)2022-04-26
中国农业科学(2021年23期)2022-01-14
农村科学实验(2021年27期)2021-11-12
医药前沿(2020年28期)2020-12-02
中国瓜菜(2020年8期)2020-09-26
儿童故事画报·发现号趣味百科(2019年9期)2019-02-02
环球时报(2019-01-03)2019-01-03
百科知识(2016年22期)2016-12-24
中国烟草学报(2012年5期)2012-04-12
