
水稻高温敏感侧根缺失突变体k209的鉴定和基因定位
2018-12-05 03:54:40林静霞王毅哲梁人尹丁沃娜朱世华郑文娟
西北植物学报 2018年10期
林静霞,王毅哲,梁人尹,刘 烨,丁沃娜,朱世华,郑文娟*
(1 宁波大学 海洋学院,浙江宁波 315211;2 宁波大学 科学技术学院,浙江宁波 315212)
根系是水稻吸收水分、养分、固定和支持植株的器官,发达的根系对植株维持正常的生理功能及抵抗不良环境具有重要的作用。侧根作为水稻胚后发育的重要器官,在水稻根构型的建立中起着重要的作用,它能够大幅度增加根系的吸收面积[1]。水稻的侧根发生源自临近原生韧皮部的中柱鞘细胞的分裂,研究表明侧根发生的位置并不完全由遗传因素决定,也受外界环境条件的影响。侧根的数量、分布及发育状况的好坏直接影响其对土壤中养分的吸收,从而改变植株在各种环境下的生长状况,最终影响作物产量以及质量[2]。现阶段对水稻的侧根发育机制了解还很少,目前已经报道了一些水稻侧根发育相关突变体[3],但经过功能鉴定的主要是几个水稻生长素途径相关蛋白,如OsAUX1[4]、OsIAA3[5]、OsIAA11[6]、OsIAA13[7]、OsIAA23[8]和OsCYP2[9]等,整体上人们对水稻侧根发生、发育的认识还不是很清楚。
温度是影响水稻生长发育的最重要因素之一。目前,高温已经成为水稻产量的主要限制因素,随着全球温室效应的加剧,高温胁迫危害水稻生产的问题愈发突出[10-11]。温度过高或过低都会抑制水稻生长发育。高温对水稻的影响不仅限于营养生长期,在其生殖期、花期都会产生不同程度的影响,而这些影响最终会造成株系不育[12]。温度对水稻根系的影响,从作用程度上讲,不及肥料与水分,但从功能上讲,却不容忽视。根际温度影响着根系的生长发育、形态结构及根中各种代谢过程。温度对水稻根系生长的影响主要表现在生长前期根系的形成和生长后期根系的衰竭。
k209是从EMS诱变的籼稻Kasalath突变体库中筛选而来的一个对高温敏感的侧根缺失突变体。该突变体在正常条件下的株高、主根和不定根的表型与野生型Kasalath基本相同,但侧根的数目与长度较野生型均有减少;在高温条件下则与野生型有着显著的区别,主要表现为地上部矮化,主根和不定根的伸长受到不同程度的抑制,最明显的性状表现为侧根完全缺失。本研究在突变体k209表型鉴定和遗传分析的基础上进行了基因定位,最终将K209定位在4号染色体上,为该基因的克隆和水稻侧根发育的分子机制研究奠定了基础。
1 材料和方法
1.1 水稻材料与培养条件
水稻高温敏感侧根缺失突变体k209是由籼稻Kasalath经EMS诱变后筛选得到。正常培养条件:光照12 h,相对湿度70%,白天32 ℃,夜晚22 ℃;高温培养条件:光照12 h,相对湿度70%,34 ℃恒温培养。水稻培养液的配方参照Yoshida等[13]。
1.2 表型分析
将野生型Kasalath和突变体k209的种子用蒸馏水冲洗干净,以0.6%稀 HNO3破休眠处理16 h,37 ℃暗处催芽约2 d至露白。将露白的种子播于水稻培养液浮着的尼龙网纱上面,在正常和高温条件下培养7 d后,用照相机(Nikon D70s)拍摄幼苗全株表型和根部性状,用标尺测量株高、主根长和不定根长,用体式镜(Leica MZ95,Germany)对根茎结合部、主根中段和根尖进行拍摄。对7 d苗龄的野生型Kasalath和突变体k209的主根进行亚甲基蓝染色,并用体式镜拍摄观察侧根原基。样本统计数为20株,3次重复。
1.3 遗传分析和定位群体的构建
用纯合突变体k209作为母本,野生型kasalath为父本杂交获得F1个体。F1种子在34 ℃条件下恒温生长7 d观察其侧根表型;F1自花授粉获得F2群体,随机选取F2群体的种子在34 ℃条件下生长7 d,统计野生型表型和突变体表型的分离比并进行χ2检验分析。突变体k209与粳稻Nipponbare杂交,F1自交获得F2群体,其在34 ℃高温条件下生长7 d后分离出来的侧根缺失植株用于基因定位。
1.4 分子标记分析
1.4.1分子标记的选择和设计用于基因定位的分子标记来自已公布的SSR序列(http://www.gramene.org/),每条染色体按照20 cM的遗传距离较均匀合理地选择约10个SSR标记进行粗定位。根据已公布的粳稻Nipponbare全基因组序列信息RGP(http://rgp.dna.affrc.go.jp/E/toppage. html)和中国华大基因研究中心提供的籼稻品种9311全基因组序列比对,在有差异的序列两侧设计引物,检测后选取在亲本间条带有差异的InDel标记对K209进行精细定位。InDel和SSR标记引物均由上海生工生物工程有限公司合成。
1.4.2PCR扩增采用简易TPS法[14]从水稻叶片中提取亲本、F1和F2分离群体的DNA。利用PCR扩增筛选的分子标记,具体的扩增体系是:1 μL模板DNA、1 μL 10×PCR buffer、1.2 μL 25 mmol/L MgCl2、0.3 μL 2.5 mmol/L dNTP、0.3 μL 10 μmol/L上下游引物、0.1 μL 5 U/mLTaqDNA聚合酶、5.8 μL超纯水。PCR反应条件是:94 ℃预变性4 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,34个循环;72 ℃延伸5 min。
配制聚丙烯酰胺凝胶,待胶凝固后,用3 μL PCR产物点样,开始电泳跑胶。跑完PAGE胶后用0.1% AgNO3银染10 min,清水洗净后,再倒入400 mL显色液显色,直至胶呈现黑色条带。
2 结果与分析
2.1 突变体的表型鉴定
两种温度环境下培养的野生型Kasalath和突变体k209的7 d幼苗表型观察发现,k209在正常培养条件下,株高、主根长、不定根长均与野生型的基本相同(图1, a、c),但侧根数目变少,只有野生型的64.19%(表1);而在高温培养条件下,k209出现了明显的突变性状,主要表现为侧根缺失,此外幼苗株高变矮,为野生型的57.18%;主根和不定根的长度都变短,分别只有野生型的55.46%和44.07%,不定根数与野生型相比有所增加(图1, b、d;表1)。在体式镜下观察根部各个部位的表型可知,正常培养条件下突变体k209的侧根长度明显变短,只有野生型的41.12%(图2和表1);在高温条件下,k209的侧根完全缺失(图2)。
2.2 突变体侧根原基发生分析
为了了解突变体k209高温侧根缺失的原因,对7 d苗龄的野生型Kasalath和突变体k209的幼苗主根进行了亚甲基蓝染色观察。结果发现,正常条件下,野生型和k209幼苗主根上均可以观察到侧根原基以及肉眼可见的侧根,k209的侧根原基数约为Kasalath的79.28%;在高温条件下,Kasalath和k209幼苗主根上均可以观察到侧根原基,但k209侧根原基数目明显少于Kasalath,约为Kasalath的58.03%,且不能突破表皮长出侧根(图3和表2)。
2.3 遗传分析


a.常温培养下整株表型;b.高温下整株表型;c.常温培养下根系表型;d.高温下根系表型。Bars=2 cm图1 7 d苗龄的野生型Kasalath与突变体k209的表型a. Seedlings under normal temperature; b. Seedlings under high temperature; c. Roots under normal temperature; d. Roots under high temperature. Bars=2 cmFig.1 Phenotypic characteristics of 7-day-old wild type Kasalath and k209 mutant
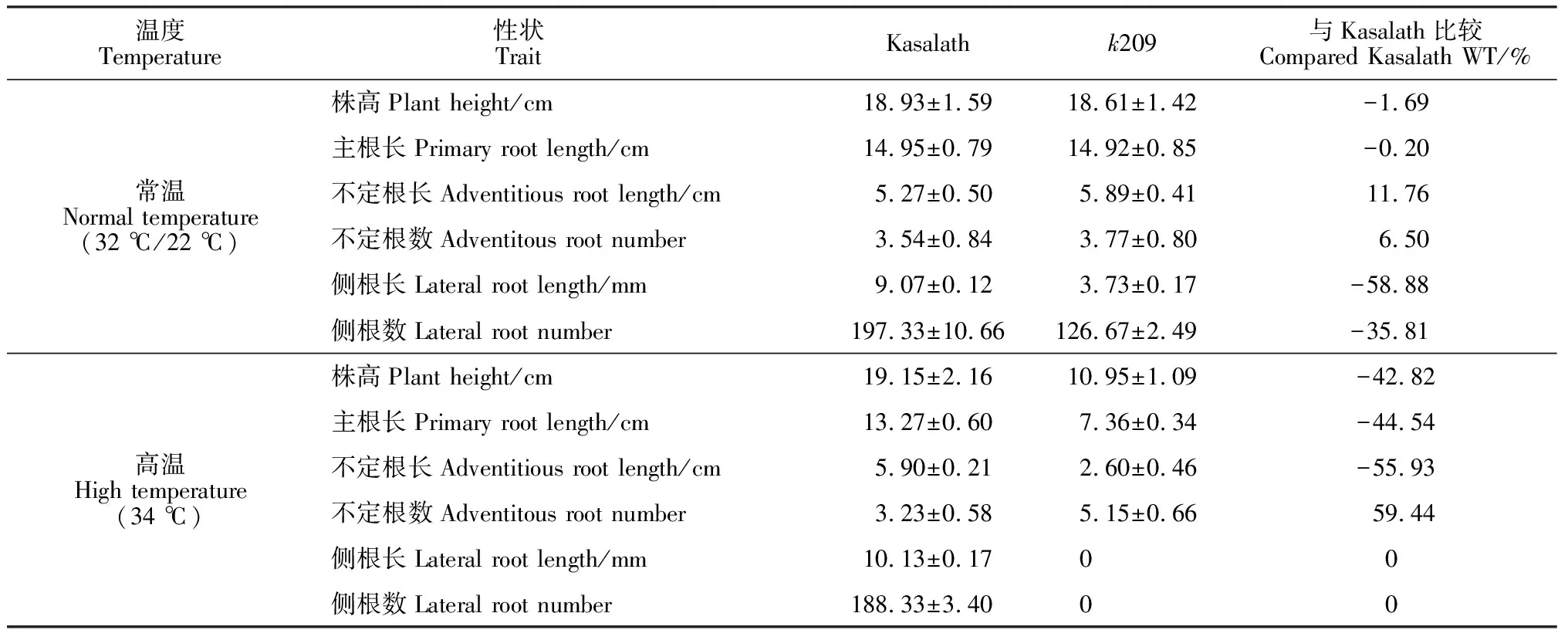
温度Temperature性状 Trait Kasalathk209与Kasalath比较 Compared Kasalath WT/%常温Normal temperature(32 ℃/22 ℃)株高Plant height/cm18.93±1.5918.61±1.42-1.69主根长Primary root length/cm14.95±0.7914.92±0.85-0.20不定根长Adventitious root length/cm5.27±0.505.89±0.4111.76不定根数Adventitous root number3.54±0.843.77±0.806.50侧根长Lateral root length/mm9.07±0.123.73±0.17-58.88侧根数Lateral root number197.33±10.66126.67±2.49-35.81高温High temperature(34 ℃)株高Plant height/cm19.15±2.1610.95±1.09-42.82主根长Primary root length/cm13.27±0.607.36±0.34-44.54不定根长Adventitious root length/cm5.90±0.212.60±0.46-55.93不定根数Adventitous root number3.23±0.585.15±0.6659.44侧根长Lateral root length/mm10.13±0.1700侧根数Lateral root number188.33±3.4000
注:不定根长为最长3根不定根长度的平均值;侧根长为最长3根侧根长度的平均值
Note:Adventitious root length is the average length of three longest adventitious roots; Lateral root length is the average length of three longest lateral roots
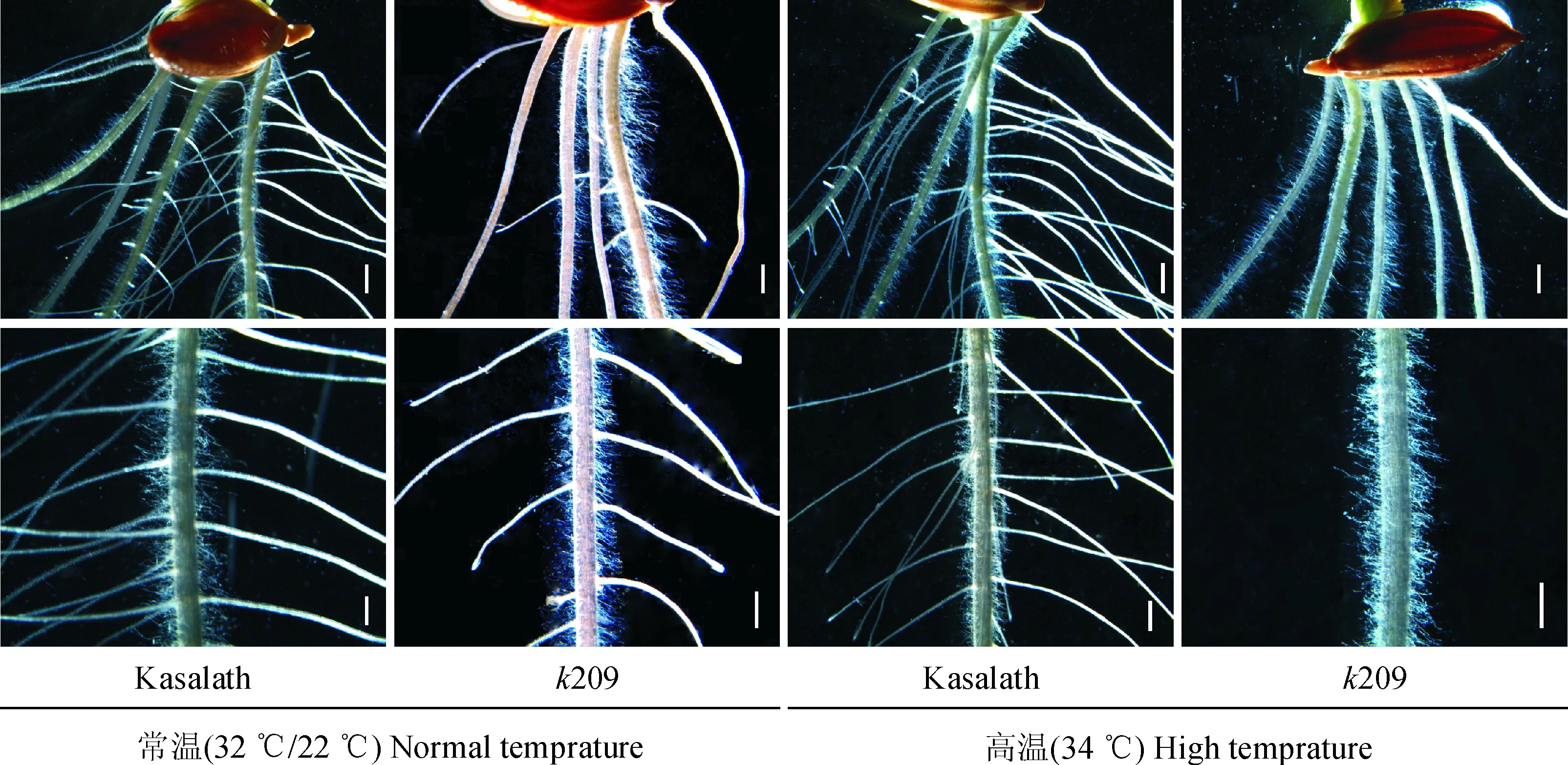
图2 7 d苗龄的野生型Kasalath与突变体k209的主根体视镜照 (Bars=1 mm)Fig.2 The primary root of the 7-day-old wild type Kasalath and k209 mutant under stereoscope (Bars=1 mm)
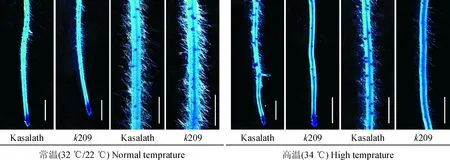
图3 7 d苗龄的野生型Kasalath和突变体k209主根侧根原基的亚甲基蓝染色Fig.3 Lateral root primordia stained with methylene blue on primary roots of 7-day-old wild type Kasalath and k209 mutant
2.4 突变体基因定位
选用粳稻Nipponbare和突变体杂交获得的F2群体作为定位群体,提取30株F2侧根缺失突变株的DNA,然后吸取等量DNA至离心管中构建一个突变池。以突变池为模板,Kasalath、Nipponbare和F1为对照,用181对现有的SSR序列引物进行PCR扩增和聚丙烯酰胺凝胶显色。结果发现在4号染色体上的物理位置为11 471 kb的分子标记RM16604处混合样品基因型为野生型Kasalath,解包检测结果为30个群体重组为零(图4),说明RM16604可能与K209基因连锁。
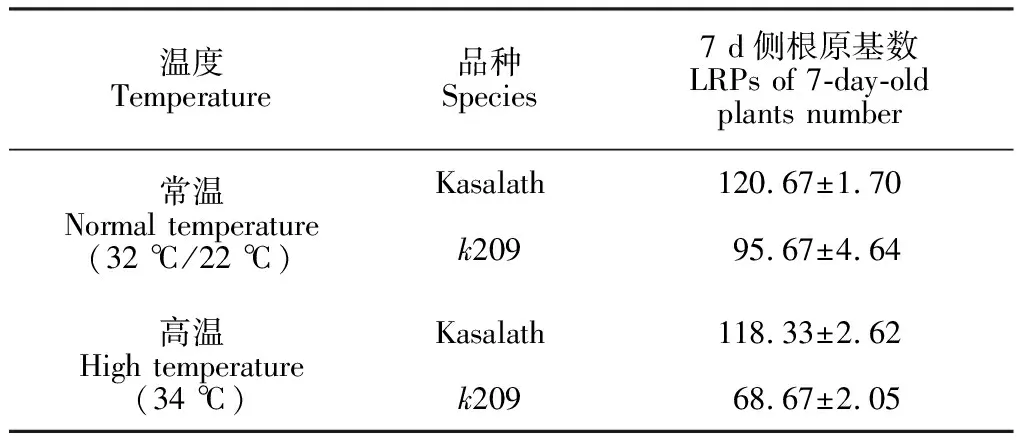
表2 不同温度条件下7 d苗龄的野生型Kasalath和
注:表中数据取3个不同主根上的侧根原基数的平均值
Note: The average number of the LRPs of three primary roots
选用RM16604附近的SSR引物和设计新的InDel分子标记进行解包分析(表4),同时不断扩大F2突变群体进行分析,RM16604和11408K分子标记处重组子数目为零。在7522K和11524K处分别检测到重组子(图5),最终将K209基因定位在标记7522K和11524K之间,物理距离大约为4 002 kb,着丝粒正好位于此区间内(图6)。
3 讨 论
植物侧根形成是根系发育的关键因素,受多方面的影响。与地上部分侧枝的形成相比,侧根形成具有更多的不确定性[15]。现阶段对禾谷类作物侧根发育的研究还较为滞后,这在一定程度上给开展以根系优良性状为目标的作物育种等实际工作造成了困难。因此,对于水稻等禾谷类作物侧根发生发育的过程和调控机理的研究极为重要。

表3 突变体k209的遗传分析
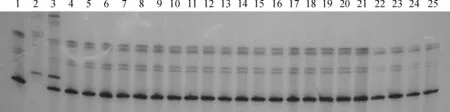
1.Kasalath;2. Nipponbare;3. F1(k209×Nipponbare);4~25. F2侧根缺失单株图4 与K209基因连锁的SSR标记RM16604的扩增结果1. Kasalath; 2. Nipponbare; 3. F1(k209×Nipponbare); 4-25. Lateral rootless individuals in the F2 populationFig.4 Amplification of the SSR marker RM16604 linked with the K209 gene
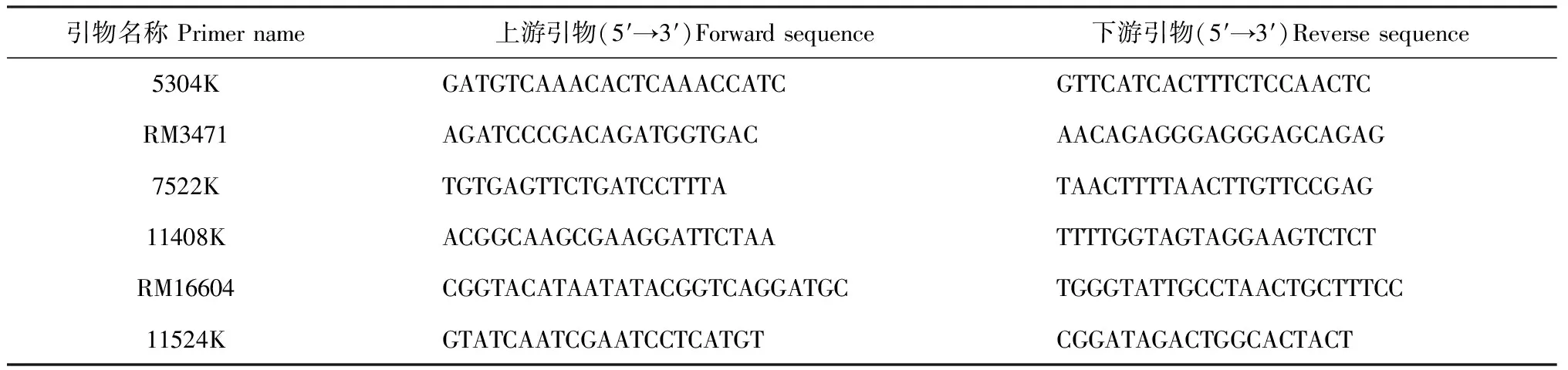
引物名称 Primer name上游引物(5′→3′)Forward sequence下游引物(5′→3′)Reverse sequence5304KGATGTCAAACACTCAAACCATCGTTCATCACTTTCTCCAACTCRM3471AGATCCCGACAGATGGTGACAACAGAGGGAGGGAGCAGAG7522KTGTGAGTTCTGATCCTTTATAACTTTTAACTTGTTCCGAG11408KACGGCAAGCGAAGGATTCTAATTTTGGTAGTAGGAAGTCTCTRM16604CGGTACATAATATACGGTCAGGATGCTGGGTATTGCCTAACTGCTTTCC11524KGTATCAATCGAATCCTCATGTCGGATAGACTGGCACTACT
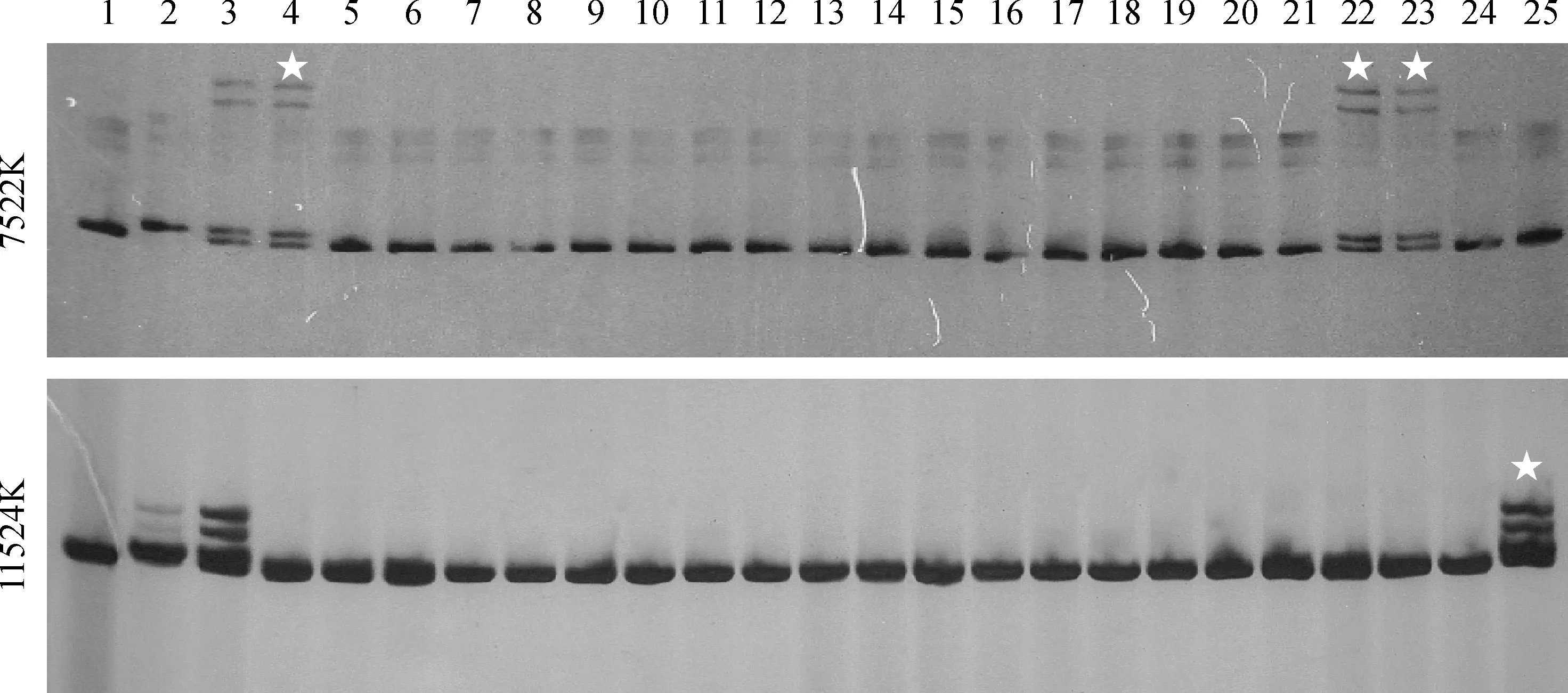
1.Kasalath;2. Nipponbare;3. F1(k209×Nipponbare);4~25. F2侧根缺失单株;☆.重组单株图5 标记7522K和11524K在Nipponbare/k209 F2群体中分离的部分扩增结果1.Kasalath; 2. Nipponbare; 3. F1(k209×Nipponbare); 4-25. Lateral root loss individuals in F2 population; ☆.RecombinantFig.5 Segregation of markers 7522K and 11524K in partial individuals of the Nipponbare/k209 F2 population
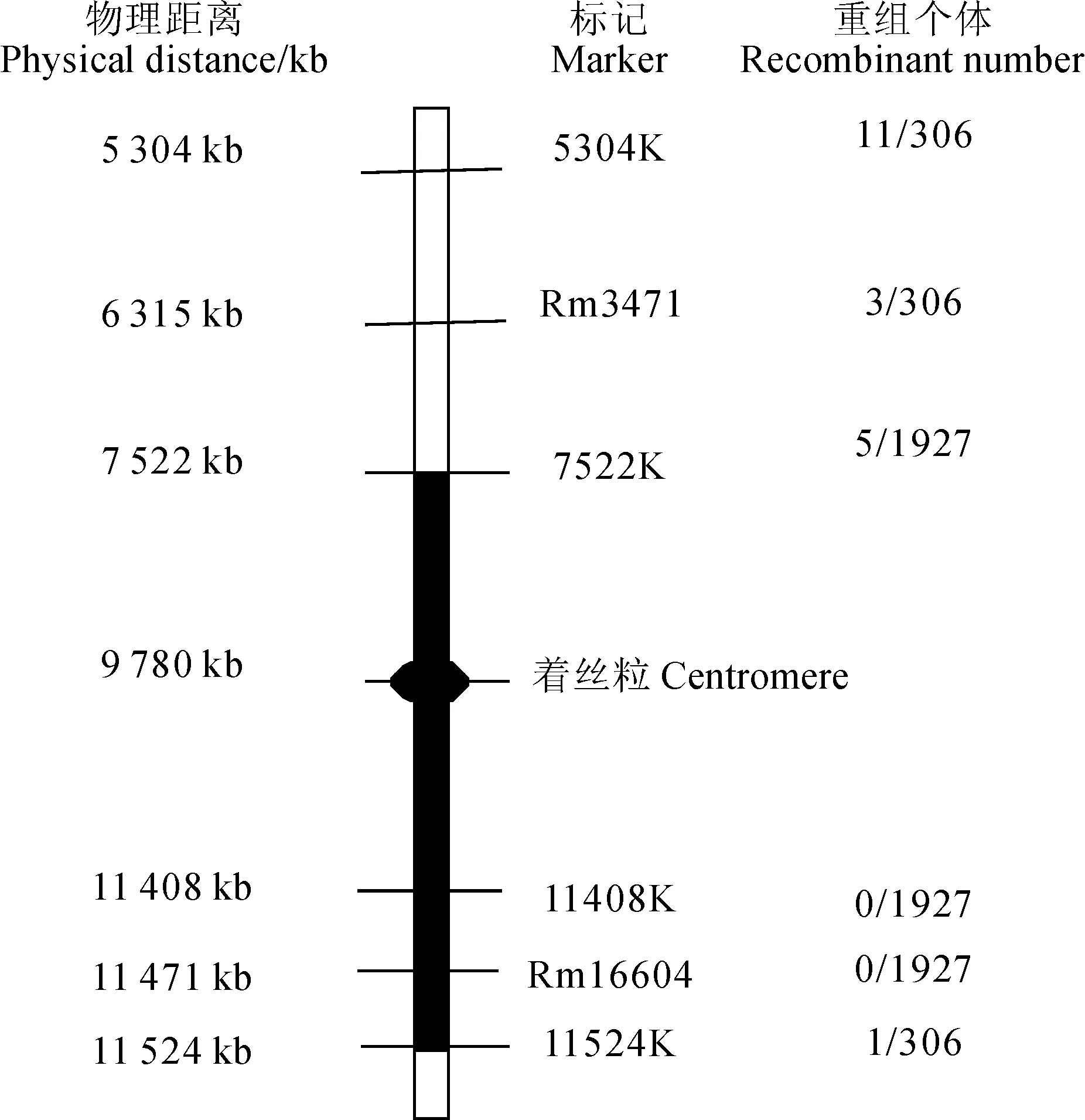
图6 K209基因在水稻4号染色体上的分子定位示意图Fig.6 Schematic diagram of the K209 gene mapped on rice chromosome 4
侧根的发生大致可以分为起始、原基形成、分生组织形成以及活化等几个关键时期[16],内在遗传调控因子和外在环境因素共同决定谷物中侧根的形成。近年来发现了一些与水稻侧根生长发育相关的突变体,它们主要表现在侧根伸长受限制以及侧根密度降低。在大多数情况下,侧根发生缺失的水稻突变体中其他类型的根也会表现出异常发育,这恰恰体现了谷类根系结构中的复杂调控机制。本研究中的侧根缺失突变体k209在正常培养温度(昼32 ℃/夜22 ℃)下的株高、主根长、不定根长及不定根数与野生型相比差异不大,但侧根的数目与长度均有减少;在高温(34 ℃恒温)条件下突变体幼苗表现为侧根完全缺失,主根和不定根的长度变短,且株高变矮。通过亚甲基蓝染色结果分析表明,在正常培养条件下,野生型和突变体k209的7 d苗龄主根上均能观测到一定数量的侧根原基以及突破表皮形成的侧根;在高温条件下,野生型和k209幼苗主根上均可以观察到侧根原基,但是k209的侧根原基数目明显少于野生型,且不能突破表皮长出侧根。可见,高温对k209的侧根原基的发生和发育都有影响。
已有研究表明,生长素对侧根发生发育的调控非常关键。在水稻中,编码Aux/IAA蛋白的OsAUX1[4]、OsIAA3[5]、OsIAA11[6]、OsIAA13[7]和OsIAA23[8]基因突变都导致了其对应的突变体侧根发生缺陷。OsAUX1基因位于1号染色体上,它参与生长素极性运输,在调控生长素介导的水稻侧根起始中发挥功能,aux1突变体主根变长,侧根减少;OsIAA3基因位于12号染色体上,突变体iaa3表现在侧根伸长受到抑制以及密度减少;OsIAA11基因位于3号染色体上,iaa11的侧根原基萌发受到抑制;OsIAA13基因位于3号染色体上,iaa13突变体的侧根密度降低;OsIAA23基因位于6号染色体上,与突变体iaa3表型类似,iaa23的侧根伸长也受到抑制同时密度减少。此外,编码亲环素蛋白的OsCYP2/LRT2基因参与Aux/IAA蛋白降解从而调控侧根发生,OsCYP2/LRT2基因位于2号染色体上,突变体cyp2和lrt2表现为对生长素不敏感,侧根缺失[17-18]。
近年来在水稻中已筛选到一些与温度相关的突变体,但和温度相关的水稻侧根突变体的报道还很少,与k209具有类似发生机制的突变体主要有hts1[19]和orc3[20]。对突变体hts1的初步研究表明,26 ℃时突变体hts1的侧根表型正常,28~32 ℃时表现为少侧根,34 ℃以上表现为无侧根以及不定根数目显著增加,目前还没有该基因的定位报道。OsORC3基因位于10号染色体上,突变体orc3在26 ℃培养下侧根正常发育,而在34 ℃高温环境下表现为侧根缺失,亚甲基蓝实验结果表明高温条件下突变体orc3的侧根原基发育正常,但最终无法突破表皮形成侧根。
本研究中的水稻高温敏感侧根缺失突变体k209属于隐性单基因突变。突变基因被定位在4号染色体的InDel标记7522K和11524K之间,目前该区间还没有侧根发育相关基因的报道,表明K209基因是一个调控侧根发育的新基因。因此该基因的克隆和功能的深入研究有助于进一步揭示植物侧根发育的分子机制。
猜你喜欢
作物研究(2022年1期)2022-11-27 23:34:38
作物学报(2022年10期)2022-07-21 03:13:58
农业现代化研究(2022年3期)2022-06-29 09:31:54
广西植物(2022年5期)2022-06-18 08:50:41
文山学院学报(2022年2期)2022-05-27 05:45:34
课外生活·趣知识(2022年5期)2022-05-14 16:26:29
现代农业研究(2021年10期)2021-11-02 14:40:24
天然产物研究与开发(2018年4期)2018-05-07 06:48:03
河北果树(2016年2期)2016-08-12 07:03:37
植物营养与肥料学报(2011年4期)2011-10-26 02:44:56
