
西藏喜马拉雅紫茉莉野生居群果实形态变异
2018-12-05 03:54汪书丽孙凤荣吉哈利
西北植物学报 2018年10期
汪书丽,罗 建,孙凤荣,吉哈利
(1 西藏农牧学院 高原生态研究所,西藏林芝 860000;2 西藏高原森林生态教育部重点实验室,西藏林芝 860000;3 西藏林芝高山森林生态系统国家野外科学观测研究站,西藏林芝 860000;4 西藏农牧学院,西藏林芝 860000)
喜马拉雅紫茉莉[Mirabilishimalaica(Edgew.) Heimerl]属于紫茉莉科(Nyctaginaceae)紫茉莉属(MirabilisL.)植物[1]。它和变种中华紫茉莉(M.himalaicavar.chinensisHeimerl)是藏药“喜马拉雅紫茉莉”(巴朱)的正品原植物[2]。以干燥根入药,被称为传统藏药的“五根”之一,主要出现在具有滋补作用的藏成药配方中,是西藏的Ⅰ级濒危藏药材之一[3]。作为一种重要的藏药材,关于其研究多数集中在化学成分和药理[4]等方面,而关于其果实方面的研究较少,主要在组织培养[5]、化感作用[6]、瘦果粘液[7]、萌发特性[3]等方面。关于喜马拉雅紫茉莉的果实形态,唐昌林[8]将紫茉莉科果实类型描述为瘦果状掺花果,而卢骁等[7]描述为瘦果。兰小中等[3]对西藏芒康县、朗县、尼木县、桑日县的喜马拉雅紫茉莉种子(掺花果)的质量和萌发特性进行了研究,并对4个县的种子(掺花果)测量了长、宽和千粒重,并进行了简单形态描述。然而,较广泛地对西藏喜马拉雅紫茉莉居群展开较全面的果实形态变异方面的分析还未见报道。掌握果实表型变异规律,是进行果实鉴别、种源区划等工作的前提和基础,为种源试验采样和试验设计提供重要依据,同时也是遗传育种的基础[9]。本研究对西藏喜马拉雅紫茉莉(包括其变种)16个种源的野生种质资源的果实表型变异进行广泛的采样和分析,讨论其种源间的果实形态差异,为进一步研究其遗传多样性和种质选育等提供基础资料。
1 材料和方法
1.1 野外调查
该研究于2014~2015年9~11月进行,分别对对西藏4个市、12个县、16个不同分布地点的喜马拉雅紫茉莉居群进行野外调查和取样,各居群的地理位置信息见表1。
1.2 性状测量
每个居群选取20~25个植株(TNC1,TNC2分别取12和15个植株),16个居群共计选取351个植株(表1)。每个植株随机选取3个果实,野外用游标卡尺测量其总苞筒长、总苞裂片宽和总苞裂片长,精度为0.01 mm。之后采集其果实装入信封并编号带回实验室。
室内处理干净每个植株的所有果实,去掉总苞。每个植株随机选取3粒健康掺花果,测量其长和宽,掺花果的形状用果长/果宽[10]表示;掺花果的形态由Nikon SMZ18体式显微镜拍摄,放大10倍;放大镜下观察果实纹路,表面无明显棱,具明显凸凹不平的嵴[3]的掺花果,赋值为1,表面有很明显的棱[7]的掺花果,赋值为2。由于喜马拉雅紫茉莉掺花果被虫子啃食非常严重,每个植株选取1 000粒称重非常困难,因此果实的重量选取百粒重指标计量,每个植株选取100粒掺花果,用电子天平称量,精确到0.01 g。
1.3 数据处理
本研究共选取 8个果实性状(表 2)。每个植株除百粒重外,其他7个性状均取3个果实的平均值代表植株的各个性状。同一个居群的所有单株果实性状的平均值代表居群性状;不同居群和总体果实性状的平均值和标准差用Excel 2010统计,变异系数(variation coefficient)用标准差/平均值表示[12];单因素方差分析、主成分分析、聚类分析和相关性分析均用SPSS19.0进行。其中,果实性状指标在居群间形态差异程度采用单因素方差分析(One way ANOVA)进行测定;主成分分析(PCA)用以确定造成居群间表型变异的主要性状;聚类分析采用离差平方和法(Ward),基于8个性状对16个居群进行聚类;相关性分析采用Pearson相关性系数分析各性状之间、各性状和地理因子之间的相关性。
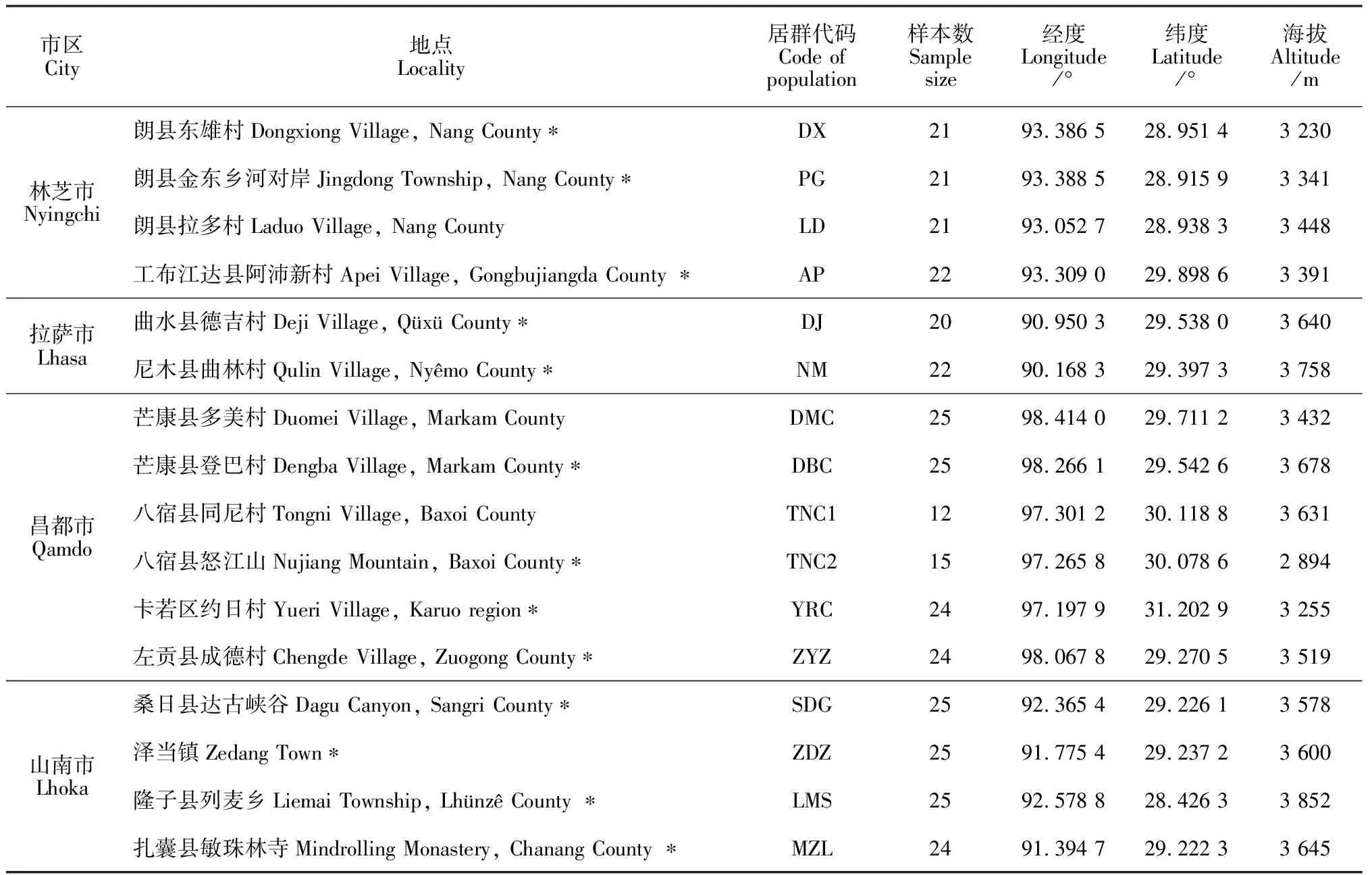
表1 喜马拉雅紫茉莉居群采集地点概况
注:*标注地点的经纬度和海拔在吉哈利[11]中引用过,此处对经纬度进行转换,并更正了个别数据
Notes: Locations marked with “*” have been cited in Ji[11]. Longitude and latitude data are transformed, and a few environmental data are corrected
2 结果与分析
2.1 不同种源喜马拉雅紫茉莉的果实性状
喜马拉雅紫茉莉16个居群主要的果实性状特征祥见图1和表2。由表2可以看出,8个果实性状的平均值在不同居群存在差异:八宿怒江山TNC2居群的总苞筒长、总苞裂片长和宽、果长、果形、百粒重均最小。虽然其果宽为(2.41±0.14)mm,稍大于最小值LMS的(2.37±0.16)mm,但综合其果长和果宽,其掺花果仍然最小;其他居群未在各个性状上表现出规律性:AP居群的总苞筒长最大、果形长宽比例最大。DJ居群的总苞裂片长和宽均最大。SDG居群的果长、果宽和百粒重最大;从果实纹路上看,各个居群均具有两种类型的掺花果,只是2种类型的果实比例稍有差异。TNC1居群值最大(1.92 ± 0.21),说明B型掺花果多于A型。ZYZ居群值最小(1.07 ± 0.17),说明A型掺花果多于B型。
各居群的不同性状的变异系数变化也存在差异:9个居群均以质量性状果纹路的变异系数最大,少数居群以百粒重或总苞裂片宽变异最大;除了LD居群的总苞筒长变异最小,多数居群以果宽或果长变异最小。将351个植株做为一个整体计算,总体上看,各性状的变异系数在8.58%~25.57%之间,总苞裂片宽变异最大,而果宽变异最小。
各个性状的单因素方差分析F值在12.180~134.761之间,均表现为极显著,说明各性状在不同居群间存在明显差异。8个形态指标在居群间的差异程度依次为:总苞筒长>总苞裂片宽>总苞裂片长>果长>果宽>百粒重>果纹路>果形。
2.2 主成分分析
按照最小特征根大于1.0的原则提取3个主成分(表3,4),其累计贡献率达81.683%,所以这3个主成分可以代替原始因子所代表的全部信息。第1主成分的特征值贡献率为48.376%,对其作用最大的性状主要为果长(0.834),其次为总苞裂片长(0.828)和总苞裂片宽(0.816);第2主成分特征值贡献率为20.091%,对其作用最大的性状主要为果形(0.865);第3主成分特征值贡献率为13.216%,对其作用最大的性状主要为果纹路(0.432)。以上分析可以得知,贡献率最大的第1主成分中的果长、总苞裂片长和总苞裂片宽的负荷均大于80%,说明果长和总苞裂片大小是造成各居群果实表型差异的主要因素。

居群代码同表1图1 不同居群喜马拉雅紫茉莉的果实形态Code of population same as Table 1Fig.1 Morphological characteristics in different populations of M.himalaica

居群代码Code of populationITL/mmILW/mmILL/mmFL/mmFW/mmFSFPFEW/gDX4.89±0.33(6.75)2.94±0.34(11.56)3.19±0.33(10.34)4.26±0.31(7.28)2.56±0.15(5.86)1.67±0.15(8.98)1.62±0.43(26.54)1.53±0.30(19.61)PG5.55±0.43(7.75)3.71±0.43(11.59)3.85±0.53(13.77)4.47±0.54(12.08)2.82±0.19(6.74)1.59±0.12(7.55)1.52±0.45(29.61)1.96±0.38(19.39)LD4.79±0.34(7.10)2.69±0.25(9.29)2.82±0.22(7.80)4.04±0.34(8.42)2.41±0.20(8.30)1.68±0.13(7.74)1.87±0.22(11.76)1.55±0.37(23.87)AP8.71±0.76(8.73)3.84±0.53(13.80)4.57±0.62(13.57)4.82±0.38(7.88)2.60±0.18(6.92)1.87±0.15(8.02)1.15±0.22(19.13)1.79±0.29(16.20)DJ6.00±0.38(6.33)4.50±0.30(6.67)4.68±0.36(7.69)4.82±0.25(5.19)2.84±0.16(5.63)1.72±0.12(6.98)1.38±0.25(18.12)2.02±0.26(12.87)NM8.56±0.65(7.59)3.85±0.30(7.79)4.43±0.36(8.13)4.86±0.26(5.35)2.80±0.23(8.21)1.75±0.17(9.71)1.47±0.29(19.73)1.76±0.60(34.09)DMC4.92±0.52(10.57)2.84±0.39(13.73)3.39±0.36(10.62)4.38±0.40(9.13)2.72±0.18(6.62)1.62±0.19(11.73)1.39±0.34(24.46)1.78±0.22(12.36)DBC5.20±0.64(12.31)2.85±0.40(14.04)3.63±0.50(13.77)4.51±0.24(5.32)2.77±0.16(5.78)1.64±0.11(6.71)1.39±0.27(19.42)1.97±0.17(8.63)TNC14.23±0.66(15.60)2.24±0.39(17.41)3.19±0.56(17.55)4.39±0.30(6.83)2.57±0.19(7.39)1.72±0.17(9.88)1.92±0.21(10.94)1.65±0.31(18.79)TNC23.84±0.59(15.36)1.76±0.31(17.61)2.63±0.35(13.31)3.66±0.29(7.92)2.41±0.14(5.81)1.52±0.11(7.24)1.78±0.24(13.48)1.25±0.16(12.80)YRC5.15±0.43(8.35)2.38±0.33(13.87)3.33±0.37(11.11)4.64±0.30(6.47)2.66±0.22(8.27)1.76±0.15(8.52)1.56±0.27(17.31)1.94±0.22(11.34)ZYZ4.99±0.61(12.22)2.78±0.80(28.78)2.96±0.40(13.51)4.88±0.29(5.94)2.76±0.18(6.52)1.78±0.12(6.74)1.07±0.17(15.89)2.09±0.29(13.88)SDG6.03±0.46(7.63)3.73±0.34(9.12)4.21±0.47(11.16)5.05±0.30(5.94)2.95±0.12(4.07)1.72±0.09(5.23)1.29±0.20(15.50)2.15±0.24(11.16)ZDZ5.18±0.49(9.46)3.22±0.39(12.11)3.68±0.49(13.32)4.18±0.25(5.98)2.73±0.13(4.76)1.53±0.09(5.88)1.25±0.15(12.00)1.69±0.16(9.47)LMS4.55±0.46(10.11)2.40±0.27(11.25)3.04±0.34(11.18)4.24±0.19(4.48)2.37±0.16(6.75)1.80±0.13(7.22)1.64±0.27(16.46)1.63±0.21(12.88)MZL5.02±0.61(12.15)2.51±0.25(9.96)3.05±0.33(10.82)4.33±0.21(4.85)2.77±0.11(3.97)1.57±0.08(5.10)1.42±0.15(10.56)1.88±0.14(7.45)总体Total5.52±1.38(25.00)3.05±0.78(25.57)3.56±0.74(20.79)4.49±0.46(10.24)2.68±0.23(8.58)1.69±0.16(9.47)1.46±0.35(23.97)1.79±0.37(20.67)F134.761∗∗65.084∗∗48.588∗∗27.547∗∗21.291∗∗12.180∗∗15.596∗∗16.605∗∗
注:ITL.总苞筒长;ILW.总苞裂片宽;ILL.总苞裂片长;FL.果长;FW.果宽;FS.果形;FP.果纹路;FEW.百粒重。下同;括号内为变异系数/%;**表示 a=0.01水平上差异极显著
Note: ITL. Involucre tube length; ILW. Involucre lobe width; ILL. Involucre lobe length; FL. Fruit length; FW. Fruit width; FS. Fruit shape; FP. Fruit pattern. FEW.100 fruits weight. The same as below; The number in the bracket is the variation coefficient. **represent significance at the level of a=0.01
2.3 居群聚类分析
依据8个果实性状对喜马拉雅紫茉莉16个居群进行聚类,结果如图2所示,在平方欧氏距离10的水平上,16个居群分为两大支:分支1包括11个居群,表现为总苞筒较短(3.84~5.20 mm)、总苞裂片较小(宽1.76~3.22 mm、长2.63~3.68 mm)。

表3 喜马拉雅紫茉莉果实性状相关矩阵的特征值、贡献率和累计贡献率

表4 喜马拉雅紫茉莉8个果实性状的主成分负荷量
分支2包括AP、NM、PG、SDG和DJ共5个居群,表现为总苞筒较长(5.55~8.71 mm)、总苞裂片较大(宽3.71~4.50 mm、长3.85~4.68 mm)。聚类分析的结果与各居群的地域分布并无直接关联,两个分支分布区域有交叉:分支1各居群分布在西藏的东部和东南部,而分支2的各居群分布在东南部。
2.4 相关性分析
由表5看出:8个果实性状两两之间Pearson相关性的28个组合中,15个组合有极显著的相关性,4个有显著的相关性,说明果实性状之间在生长发育上是相互影响的。其中,受较多性状显著影响的主要有3个性状:果长与其他7个性状均存在极显著的相关性,总苞裂片长和总苞裂片宽均与除了果形外的其他6个性状存在显著或极显著的相关性;与果形存在显著相关性的性状最少,仅总苞筒长和果长与其存在极显著的正相关;果纹路和其他7个性状均表现为负相关,其中和总苞裂片宽、总苞裂片长、果长和百粒重均达到显著或极显著水平。

图2 喜马拉雅紫茉莉果实形态聚类图Fig.2 The clustering dendrogram of fruit traits of M. himalaica with squared Euclidean distance

指标IndexITLILWILLFLFWFSFPFEWITL1ILW0.736∗∗1ILL0.823∗∗ 0.922∗∗1FL0.653∗∗ 0.685∗∗ 0.720∗∗1FW0.389 0.648∗∗ 0.628∗∗0.726∗∗1FS0.511∗∗0.2770.353 0.639∗∗-0.0621FP-0.496 -0.536∗ -0.508∗-0.642∗∗-0.647-0.208 1FEW0.315 0.545∗ 0.512∗ 0.848∗∗0.832∗∗0.313-0.665∗∗1海拔Altitude0.251 0.3230.3290.4370.3050.327-0.228 0.436经度Longitude-0.483-0.591∗ -0.489 -0.177 -0.185-0.0590.101-0.046纬度Latitude0.032-0.227 0.0380.095 0.0310.0940.0670.025
注: **相关性极显著(P<0.01); *相关性显著 (P<0.05)
Note: **correlation is significant at the 0.01 level; *correlation is significant at the 0.05 level
3个地理因子与8个果实性状相关性的24个组合中,仅经度和总苞裂片宽呈显著的负相关,其他组合相关性均不显著。结果说明果实性状中仅总苞裂片宽受到地理因子经度的影响,经度越大,总苞裂片宽越小。而海拔、经度和纬度对其他性状并无明显的影响。
3 讨 论
3.1 喜马拉雅紫茉莉的果实表型变异
综合平均值、变异系数和单因素方差分析结果得出,总苞筒长、总苞裂片宽、总苞裂片长、果长、果宽、果形、果纹路和百粒重8个果实性状在喜马拉雅紫茉莉不同居群间都存在极显著差异,说明其果实性状变异在不同居群间很丰富,这在一定程度上表明喜马拉雅紫茉莉有较大的选育潜力,对其种源利用等有重要指导作用,也对其种质资源的遗传多样性的评价有一定意义[13]。主成分分析显示果长和总苞裂片大小(包括长和宽)能反映喜马拉雅紫茉莉居群果实性状的大部分信息,是造成各居群间果实表型差异的主要因素。果实表型性状的变异不仅受到内部遗传分化的影响,还与所处的生长环境密切相关[9]。调查发现,喜马拉雅紫茉莉在西藏主要分布在雅鲁藏布江、怒江、澜沧江及其支流形成的干热或者干暖河谷中,山体的隔离会造成不同居群生存的小生境。植物的形态特征是长期适应进化的结果,通过个体水平的表型可塑性实现对异质环境的适应[10,14]。喜马拉雅紫茉莉在适应外界环境的过程中发生了丰富的形态变异,也表明喜马拉雅紫茉莉对环境的适应能力很强。另外,内在的遗传因素也是造成其居群间遗传变异的主要因素之一,还需要进一步从分子水平进行深入探讨。
兰小中等[3]提及其种子(掺花果)颜色分为黑色和棕色2种,可能与成熟度或者分泌粘液的多少有关。本研究调查发现黑色和棕色掺花果之间存在颜色过渡,不易判断,反而果实表面的纹路在调查的居群中发现存在棱不明显,嵴明显和棱非常明显2种形态较易判断,因此以果纹路作为果实性状之一。
3.2 居群水平的聚类分析
本研究依据果实性状进行的聚类分析的结果并没有地域分布的规律性,2个分支的分布区域上存在交叉。形成两个果实性状数值上有一定差异的分支可能与各分支内的居群分布的小生境或遗传比较相似有关。而朗县金东乡的2个居群DX和PG虽然地理距离并不远,但由于处于同一河流不同流向位置的河谷,2个居群分别聚类在分支1和2中;据《西藏植物志》[1]记载,喜马拉雅紫茉莉原变种在西藏昌都和八宿县有分布。蔡翠萍等[12]认为喜马拉雅紫茉莉个体在雄蕊数目上的丰富变异使其种下分类缺乏有效性状支持。从本研究聚类结果上看,昌都的YRC居群和八宿县的TNC1和TNC2居群虽然均在分支1上,但并未形成独立的分支,说明文献记载原变种分布地点居群在果实形态上与其他居群并无区分,我们由此推测果实形态对喜马拉雅紫茉莉种下等级的区分分类学意义不大。
3.3 环境因子与果实表型变异的关系
除总苞裂片宽与经度呈显著的负相关外,经度、纬度和海拔对8个果实性状并不存在显著影响。这可能因为本研究选取的环境因子有限,而其他更多的环境因子如温度、降雨量、土壤成分等可能会对喜马拉雅紫茉莉的表型变异产生影响。
猜你喜欢
山东林业科技(2022年3期)2022-07-14
美食(2022年5期)2022-05-07
课外生活(小学1-3年级)(2021年8期)2021-08-28
舰船科学技术(2021年1期)2021-03-09
广西植物(2020年13期)2020-09-12
河北果树(2020年1期)2020-02-09
山东工业技术(2019年12期)2019-05-30
作文评点报·小学三、四年级(2019年41期)2019-01-03
现代园艺(2018年3期)2018-02-10
创新作文(小学版)(2018年6期)2018-01-26
