
中间锦鸡儿叶表皮微形态的经度格局及其影响因素
2018-12-05 03:45张亚杰杨九艳耿倩倩李奉时
西北植物学报 2018年10期
张亚杰,杨九艳,2*,耿倩倩,杨 欢,李奉时
(1 内蒙古大学 生态与环境学院,呼和浩特 010021; 2 蒙古高原生态学与资源利用教育部重点实验室暨内蒙古自治区草地生态学重点实验室,呼和浩特 010021)
植物的形态结构是长期适应外界环境的结果[1]。叶片作为植物的同化器官,叶片的性状尤其是叶表皮的性状[1]对气候变化响应敏感[2],并对不同的环境特征表现出了较强的适应性。
植物叶片表皮的表皮毛作为植物对干旱环境的一种适应,可以有效避免强光对植物叶片的损伤,并在气孔开放时,有效减小蒸腾[1]。而叶表皮的气孔是植物进行水分和气体交换的重要通道,气孔调节是植物抵御胁迫、适应不同环境的一种积极的策略[3]。在短时间内,气孔通过开闭在水分散失和CO2吸收间起重要的平衡作用[4],而在较长时间内,气孔则通过其分布、形态、数量及大小的变化来调控气体进出[5]。在大的时空尺度上,气孔的特性,尤其是气孔密度和气孔长度是对环境长期适应的结果[6],并且二者具有高度的协同进化趋势[7-8]。在扫描电镜下,植物叶表皮表现出不同的微形态特征,如角质层的有无,表皮毛及气孔的疏密状况,气孔大小等,这些都可以作为我们研究叶表皮对环境因子适应性的材料。
经纬度以及海拔梯度通常包含了温度、降水、光强等多种环境因子的复杂变化,是研究叶表皮微形态对不同环境因子适应性的良好场所。Korner等认为由于光照强度对气孔密度的促进作用,气孔密度随海拔的增加而增加[6]。王瑞丽等[5]的研究结果表明,在低纬度地区,植物叶片的气孔密度较高,气孔长度较短。而田苗[9]研究发现,气孔长及气孔密度随纬度变化没有明显的规律,但气孔长与最大月均温呈显著的负相关关系,气孔密度与最大月均温有显著正相关关系。
灌木是陆地生态系统的重要组成部分,它具有提高植物物种多样性、改良土壤功能、水土保持、净化环境等多种重要的生态效能[10]。在将来,土地利用变化会导致许多干旱及半干旱地区发生灌木入侵,灌木会覆盖在更多的陆地上[11]。因此在气候变化的研究中灌木值得更多的关注。
中间锦鸡儿(Caraganaintermedia)是一种优良的防风固沙灌木,对环境胁迫有较强的适应性,在内蒙古高原有较广的分布,这对研究叶表皮微形态沿经度梯度对环境因子的适应性提供了很好的材料。本研究选取内蒙古高原分布的中间锦鸡儿,在扫描电镜下观测其叶表皮特征,探讨叶表皮微形态的经度格局及其影响因素,丰富其适应进化理论,为合理利用资源提供科学依据。
1 材料和方法
1.1 研究区及样品的采集
研究区设置在内蒙古高原中间锦鸡儿的自然分布区,地理位置为108.49~113.71°E,40.48~43.84°N,年平均温度3.19~7.26 ℃,年平均降水量186.4~300.84 mm,干旱指数13.19~17.26。按照从东到西干旱指数[9]递增的趋势,在典型草原、荒漠草原和草原化荒漠设置8个中间锦鸡儿样地(图1),每个样地面积为100 m×100 m。在设定的8个样地中,每个种群(表1中ci1~ci8,与图1中1~8相对应)随机选取健康成熟的6株中间锦鸡儿,采摘枝条中部的叶片,制成干样备用。各种群采样地点见图1和表1。
1.1.1样地土壤数据来源在每个种群的采样地分别采取3个重复的土样,每一重复分为0~10 cm、10~20 cm、20~30 cm 3个土层。采集的样品带回实验室进行全氮、全磷、有机碳、pH、全碳5个土壤指标的测量,所用方法分别为半微量凯氏定氮法、钼蓝比色法、重铬酸钾外加热氧化法、pH计。土壤数据测量结果见表2。
1.1.2气象数据来源表3为8个样地年降水量、平均气温、有效积温、平均相对湿度、年日照时数5个气象因子近50年的平均值,所有数据来自中国气象科学数据共享服务网。

图1 各种群采集地分布图Fig.1 Sampling site distribution map

样地 Sampling site种群编号Population number植被类型Vegetation type土壤类型Soil type干旱指数de Martonne aridity index采集地Collecting place1ci1典型草原Typical grassland淡栗钙土Light chestnut soil13.19苏尼特左旗 Sonid Zuoqi2ci2淡栗钙土Light chestnut soil13.19苏尼特左旗 Sonid Zuoqi3ci3棕钙土Brown calcic soil15.12苏尼特右旗 Sonid Youqi4ci4荒漠草原Desert grassland淡栗钙土Light chestnut soil15.12四子王旗 Siziwangqi5ci5棕钙土Brown calcic soil14.23达茂旗 Damaoqi6ci6棕钙土Brown calcic soil15.31乌拉特中旗 Wulate Zhongqi7ci7草原化荒漠Prairie Desert棕钙土Brown calcic soil15.31乌拉特中旗 Wulate Zhongqi8ci8棕钙土Brown calcic soil17.26乌拉特前旗 Wulate Qianqi
1.2 研究方法
1.2.1叶片上下表皮特征观测及计算利用TM3030Plus型扫描电镜观察叶片表皮特征。在每个种群中选取6个单株,每个单株随机选取2片成熟健康的叶片,共计12片,随机选取其中6片用于上表皮观测,其余6片用于下表皮的观测。在每一叶片叶脉两侧各取一个视野,共12个视野,观测表皮毛数、气孔数和表皮细胞数,测定30个气孔的大小,取平均值。表皮毛密度、气孔密度、气孔指数[3,11-12]按照以下公式计算:气孔密度 = 气孔数/面积(个/mm2);表皮毛密度 = 表皮毛数/面积(个/mm2);气孔指数 = 100×气孔数/(气孔数+表皮细胞数)。
1.2.2变异系数及可塑性指数计算为指征叶表皮微形态性状的离散程度,进行变异系数的计算,变异系数越小,则性状离散程度越小;同时计算中间锦鸡儿各表皮微形态的可塑性指数,以探讨植物叶表皮主要通过哪几种性状的改变适应环境。指标计算[12]公式如下:
表型性状变异系数(cofficient of variation,CV)= 标准差/平均值
可塑性指数(plasticity index,PI)=(最大值-最小值)/最大值
1.3 数据分析
利用R3.4.2软件进行单因素方差分析,分析8个种群中叶表皮微形态指标的差异性,并用多重比较对均值间的差异进行比较。以经度为自变量,叶表皮微形态指标为因变量,利用R3.4.2软件进行简单线性回归分析,分析叶表皮微形态的经度变异格局[8]。应用SPSS19.0对叶表皮微形态指标与土壤因子、气象因子做逐步回归分析。
研究对象选择2015年8月~2016年8月于我院治疗的80例咽喉炎患者,男性42例,女性38例;最大年龄72周岁,最小16周岁,平均(41.25±5.59)岁;病程最短2天,最长12天,平均(5.21±1.25)天。纳入标准:所有患者均符合急、慢性咽喉炎诊断标准,患者基本生命体征稳定,均出现咽部灼热、干燥,咳嗽及吞咽疼痛等症状,对本探究知情且签署同意书,本次探究前5天未使用抗生素治疗。排除标准:患者合并下呼吸道感染、急性肺栓塞等呼吸道疾病,病历资料不全者,不配合本次探究者。

表2 中间锦鸡儿各样地土壤养分

表3 中间锦鸡儿各样地气象数据
2 结果与分析
2.1 不同种群叶表皮微形态
中间锦鸡儿上下表皮的表皮毛均为单细胞非腺毛(图版Ⅰ,1~8、17~24),上下表皮细胞形态均为不规则形,角质层粗糙,垂周壁式样主要为平直或拱形,气孔轴式均为不定式(图版Ⅰ,9~16、25~32),表皮均为下陷气孔,为抗旱特征。
2.1.1叶片上表皮微形态特征中间锦鸡儿上表皮气孔密度、气孔指数、表皮毛密度、气孔器长、气孔器宽(均值±标准误)分别为(302.81±11.54)个/mm2、(9.94±0.38)、(204.81±12.08)个/mm2、(11.78±0.14)μm和(5.95±0.09)μm,变化范围分别为194.29~409.58个/mm2、6.37~14.16、118.16~310.64个/mm2、10.67~13.39 μm和5.31~6.53 μm。叶片上表皮微形态性状指标中表皮毛密度的变异最大(CV=0.53),气孔器长的变异程度最小(CV=0.19)(表4和图2)。
对比8个种群,种群ci2、ci3的气孔密度最大,ci5、ci7的气孔密度最小;种群ci3的气孔指数显著大于其他种群(P<0.05),ci7的气孔指数最小;种群ci8的表皮毛密度显著大于其他种群(P<0.05),ci4的表皮毛密度显著小于其他种群(P<0.05); 种群ci4的气孔器最长,ci8的气孔器最短;而种群ci6、ci7的气孔器最宽,ci1、ci3的气孔器最窄(表4)。
2.1.2叶片下表皮微形态特征中间锦鸡儿下表皮气孔密度、气孔指数、表皮毛密度、气孔器长、气孔器宽(均值±标准误)分别为(272.83±14.23) 个/mm2、(10.43±0.50)、(187.45±13.15)个/mm2、(12.49±0.42) μm、(6.18±0.09) μm,变化范围分别为131.28~481.34个/mm2、6.15~18.91、71.88~349.62个/mm2、11.04~12.99 μm、5.37~7.06 μm。叶片上表皮微形态指标中表皮毛密度的变异最大(CV=0.63),气孔器长的变异程度最小(CV=0.21)(表5和图2)。
在8个种群中,种群ci3的气孔密度显著大于其他种群(P<0.05),ci5的气孔密度最小;种群ci3的气孔指数显著大于其他种群(P<0.05),ci7的气孔指数最小;种群ci8的表皮毛密度显著大于其他种群(P<0.05),ci3的表皮毛密度显著小于其他种群(P<0.05);种群ci8的气孔器最长,ci7的气孔器最短;而种群ci2的气孔器最宽,ci3的气孔器最窄(表5)。
对比分析中间锦鸡儿上下表皮特征发现,上表皮的气孔密度和表皮毛密度大于下表皮,而气孔指数和气孔尺度小于下表皮。

表4 中间锦鸡儿叶片上表皮微形态在不同种群的统计特征
注:同一列数据后不同小写字母表示差异显著(P<0.05)。下同
Note:Different normal letters in the same column indicate significant difference at 0.05 level. The same as below

表5 中间锦鸡儿叶片下表皮微形态在不同种群的统计特征
2.2 叶表皮微形态的经度格局
2.2.1叶片上表皮微形态的经度变异中间锦鸡儿叶片上表皮气孔密度、气孔指数表现出了相似的经度变化格局,具体表现为:随经度增加,气孔密度(图3,A)和气孔指数(图3,B)均极显著地增加(P< 0.001);表皮毛密度(图3,C)和气孔器宽度(图3,D)表现出了相反的趋势(P< 0.01),并且气孔器宽度沿经度的变化趋势较弱(R2=0.05,图3,D)。

图2 中间锦鸡儿上下表皮叶表皮微形态指标的变异系数(CV)及可塑性指数(PI)Fig.2 Variation coefficient (CV) and plasticity index (PI) of leaf epidermal micromorphology
而气孔器长度则未表现出经度变化格局。
2.2.2叶片下表皮微形态的经度变异中间锦鸡儿叶片下表皮气孔密度(图4,A)、气孔指数(图4,B)均随经度增加显著地增加;表皮毛密度(图4,C)则表现出了相反的趋势。但是气孔器长度和气孔器宽度均未表现出沿经度变化的趋势。
除了气孔器宽度,中间锦鸡儿的上下表皮表现出了一致的经度变异格局,均表现为:气孔密度与气孔指数随经度的增加而增加(P< 0.001);表皮毛密度随经度的增加而减小,并且表皮毛密度沿经度的变异较弱(P< 0.01,R2=0.12)。
2.3 中间锦鸡儿叶表皮微形态的可塑性
中间锦鸡儿叶片上下表皮的变异系数均为表皮毛密度最大、气孔器长最小(表4、表5、图2)。上下表皮的可塑性指数排序均为:表皮毛密度>气孔密度>气孔指数>气孔器宽>气孔器长(图2),可见中间锦鸡儿的叶片主要通过调节表皮毛密度及气孔密度来响应环境的变化。
2.4 叶表皮微形态经度变异的影响因素
分别以中间锦鸡儿植物的气孔密度、气孔指数、表皮毛密度、气孔器长、气孔器宽为因变量,以20个气候土壤因子(年均降水量、平均气温、有效积温、平均相对湿度、日照时数,0~10 cm、10~20 cm、20~30 cm土层的土壤全氮、全磷、全碳、有机碳及pH)为自变量做逐步回归分析,引入变量P=0.05,剔除变量P=0.1,以探究影响中间锦鸡儿叶表皮微形态变异的主要环境因素。

*P <0.05;**P<0.01; ***P <0.001。下同图3 上表皮微形态的经度变异性* P <0.05; ** P <0.01; *** P <0.001. The same as belowFig.3 Longitudinal variability of upper epidermal micromorphology
2.4.1叶片上表皮微形态经度变异的影响因素对于中间锦鸡儿的叶片上表皮,在以气孔密度和气孔指数为因变量的逐步回归分析中,均可引入变量日照时数,日照时数对气孔密度和气孔指数的贡献程度分别为89.3%、83.6%,方差分析表明这2个模型的解释率显著(P<0.05);在以表皮毛密度为因变量的逐步回归分析中,引入了变量20~30 cm全磷,它对表皮毛密度的解释程度为94.9%,方差分析模型表明达到显著水平(P<0.05);在以气孔器长为因变量的逐步回归分析中,可引入变量有效积温,它对气孔器长的解释程度为64.4%,方差分析模型表明达到显著水平(P<0.05);在以气孔器宽为因变量的逐步回归分析中,可引入变量20~30 cm全氮,它对气孔器宽的解释程度为93.5%,方差分析模型表明达到显著水平(表6)。
2.4.2叶片下表皮微形态经度变异的影响因素对于中间锦鸡儿的叶片下表皮,在以气孔密度为因变量的逐步回归分析中,可引入变量20~30 cm全碳,其对气孔密度的贡献程度为66.1%,方差分析表明这个模型的解释率显著(P<0.05);在以表皮毛密度为因变量的逐步回归分析中,引入了20~30 cm全碳和相对湿度2个变量,它们对表皮毛密度的共同解释程度为94.5%,其中20~30 cm全碳对表皮毛密度的单独解释度为78.6%,大于相对湿度的解释程度,方差分析模型表明模型达到显著水平(P<0.05);在以气孔器长为因变量的逐步回归分析中,可引入变量0~10 cm全磷及20~30 cm全磷,它们对气孔器长的共同解释程度为98.5%,0~10 cm全磷对气孔器长的单独解释度为88.6%,方差分析表明模型达到显著水平(P<0.05);而在以气孔器宽为因变量的逐步回归分析中,未引入任何变量(表7)。
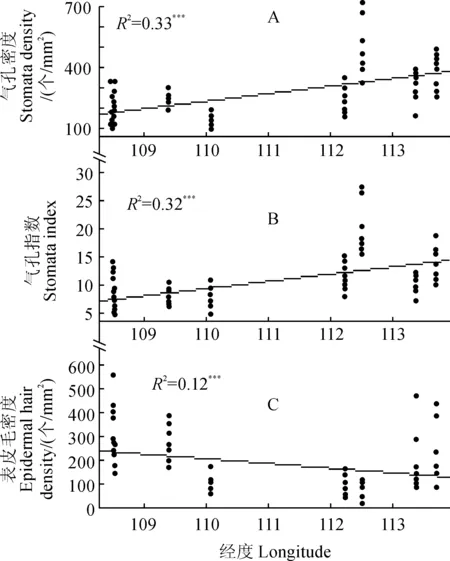
图4 下表皮微形态的经度变异性Fig.4 Longitudinal variability of lower epidermal micromorphology
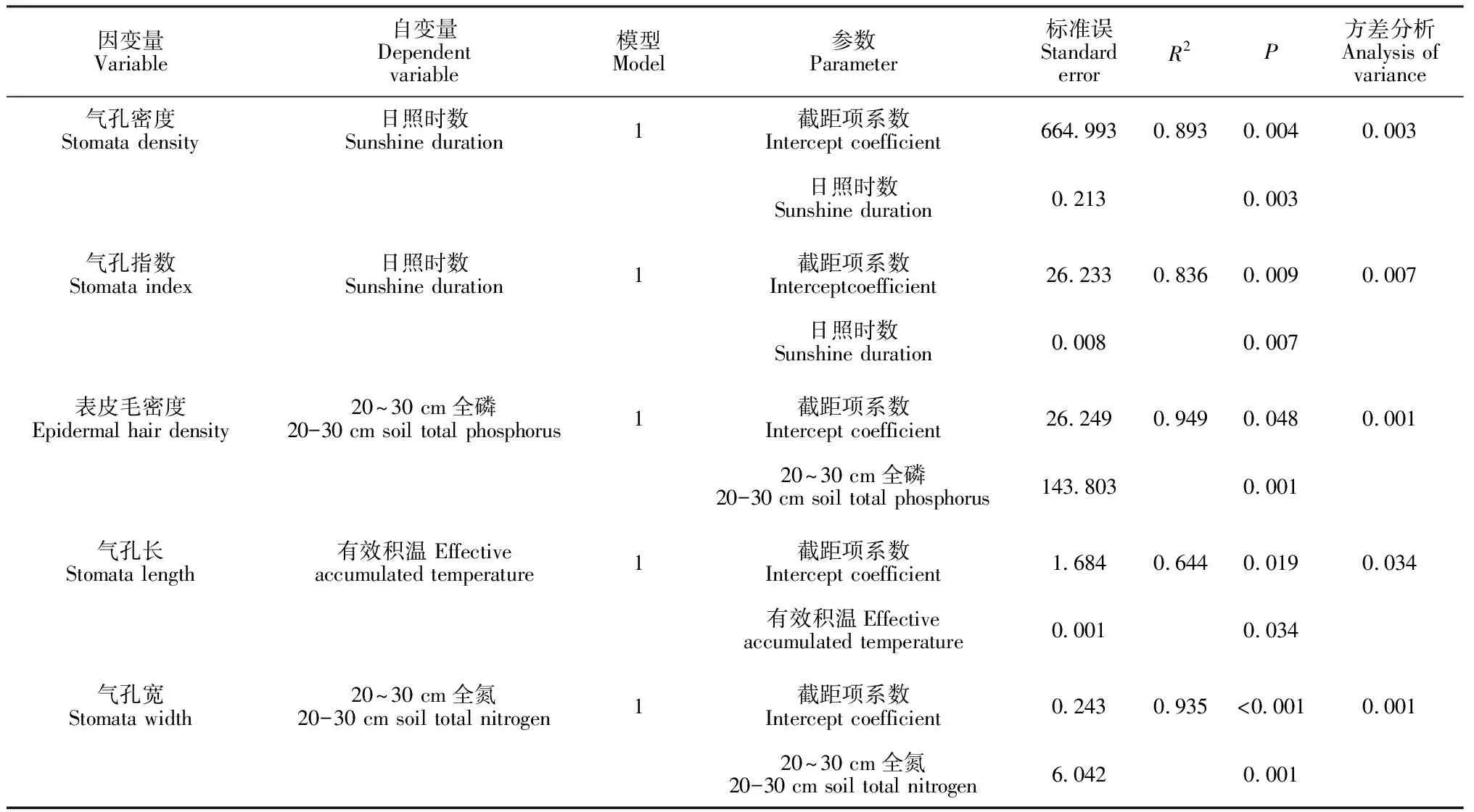
表6 中间锦鸡儿叶片上表皮微形态各指数与气象因子、土壤因子逐步回归结果

表7 中间锦鸡儿叶片下表皮微形态各指数与气象因子、土壤因子逐步回归结果
上述分析结果表明,日照时数、有效积温、相对湿度、土壤全碳、全磷、全氮含量显著影响叶片表皮的微形态特征。
3 讨 论
植物的表型变异可分为种间变异和种内变异,这是由遗传和环境因子共同决定的。其中种内变异主要是由环境因子引起的,而生态分布区及地理位置是引起居群间变异的主要因素[12]。叶片是植物在环境中暴露最多的器官,相比于植物其他营养器官(根及茎),叶片最易受到环境因子的影响[3,13],其形态结构对外界环境的变异最敏锐[14],但同时又有一定的稳定性[15-16]。叶表皮的特征随经纬度及海拔的变化呈现出不同的变异格局。随海拔的变化格局表现为:气孔密度随海拔的增加而增加[6,17]或呈现先增加后减小的趋势,气孔的孔径大小随海拔的增加而减小[18]或在不同海拔梯段呈现不同的趋势[19]。而王瑞丽等[5]研究认为物种水平上气孔密度和大小均与海拔梯度无显著关系,而在不同功能型(乔木、灌木、草本)上气孔特性与海拔梯度显示出不同变异格局。随纬度的变化格局表现为:在低纬度地区,植物叶片的气孔密度较高,气孔长度较短或气孔长度及气孔密度随纬度变化没有明显的规律[7]。而对于经度变异格局的研究较少。本实验结果表明,中间锦鸡儿的上下表皮均表现为气孔密度与气孔指数随经度的增加而增加,表皮毛密度随经度的增加而减小。
植物利用资源的能力受植物叶片性状的显著影响[20]。气孔是叶片上的重要器官,其数量和大小直接反映了光合能力的强弱,并且气孔特性(数量和大小)与植物的净初阶生产呈正相关关系[7]。气孔特征在不同植物间有较大差别,同时环境因子对气孔的发育也会产生重大影响[21],有关气孔与环境因子间关系的研究有很多不一致的结论,有研究表明气孔密度和大小主要受水热综合因子的影响,且气孔密度随年均降水量的增加以及温度的减小而减小,气孔的孔径大小则呈现相反的趋势[18-19],并且也有研究指出低温和较高的光强导致了比较高的气孔长度以及较低的气孔密度[22]。同时,从化石植物得到的气孔密度和气孔指数也表明温度波动会对气孔的数量特征产生影响[23]。本研究结果表明,在内蒙古高原从东到西分布的中间锦鸡儿,随着经度的减小即水分的减少和热量的增加,气孔密度与气孔指数下降,这样的经度格局有利于在更干旱的环境中降低水分的散失,是中间锦鸡儿应对干旱的对策之一。但是有研究评估了土壤养分(土壤氮含量)和气候因子(年均温、年降水)对气孔的数量、气孔大小这两个特征的综合影响,结果表明在大的环境梯度上,物种水平的气孔特性受到的环境因子的影响很弱[7]。在分别评估了多种不同的气候和土壤因子对上下表皮气孔特性的影响后,本实验分析结果表明:叶片上下表皮的气孔特性受不同因子的影响,叶片上表皮的气孔特性主要受气候因子的影响,其中主要受有效积温和日照时数影响;而叶片下表皮主要受深层土壤养分(0~30 cm的土壤全碳和全磷)的影响。这可能是由于上表皮在进化过程中受到的日照辐射较多而下表皮因其所处位置及角度受到一定的遮阴效果,进而叶片上表皮对气候因子表现出了较强的相关性。
表皮毛作为植物适应干旱环境的重要特征,可以有效减少阳光对植物叶表皮的直接照射[24],并可以起到反光及保温的作用[15],并减少蒸腾。气孔的数量、表皮毛的多寡等因素影响着植物在干旱环境条件下的适应性[24]。在内蒙古高原,从东到西干旱程度、日照和热量水平呈梯度增加,本实验结果表明,中间锦鸡儿的上下表皮均表现为随经度的减小表皮毛密度增加,这样的特征变化有利于中间锦鸡儿在更干热的气候环境中通过覆盖的表皮毛反射强光、降低叶温,避免表皮细胞被灼伤,同时也起到保水的作用。与下表皮相比,中间锦鸡儿叶片上表皮被有更多的表皮毛。
表型可塑性是同一物种在不同环境下产生的不同表型特性[20],与其生态适应能力密切相关[17],这是植物在没有发生遗传变异时产生的一种适应外界环境,获得有效资源,抵御胁迫的有效机制[20]。中间锦鸡儿的表皮毛密度和气孔密度具有较大的可塑性指数,表明当环境发生较大的变异时,中间锦鸡儿可以通过调节表皮毛密度、气孔密度和气孔指数适应环境变化。

图版Ⅰ 中间锦鸡儿8个种群叶表皮扫描电镜观察
1~8. 示8个种群(ci1~ci8)叶片上表皮表皮毛,× 500; 9~16. 示8个种群(ci1~ci8)叶片上表皮气孔及表皮细胞,× 3 000; 17~24. 示8个种群(ci1~ci8)叶片下表皮表皮毛,× 500/600; 25~32. 示8个种群(ci1~ci8)叶片下表皮气孔及表皮细胞,× 3 000
Plate Ⅰ Electron microscopy scanning of leaf epidermis of 8 Caragana intermedia populations
Fig.1-8. Leaf surface fur of upper epidermis of 8 populations, × 500; Fig. 9-16. Leaf stomata and cells of upper epidermis of 8 populations, × 3 000;Fig.17-24. Leaf surface fur of lower epidermis of 8 populations, × 500/600; Fig. 25-32. Leaf stomata and cells of the lower epidermis, × 3 000
猜你喜欢
种子(2022年5期)2022-06-27
干旱地区农业研究(2022年1期)2022-01-28
水土保持研究(2020年1期)2020-04-27
装备制造技术(2019年12期)2019-12-25
学校教育研究(2019年21期)2019-11-11
科技创新与品牌(2019年12期)2019-02-06
农民致富之友(2018年8期)2018-06-27
西藏农业科技(2018年4期)2018-04-25
天文爱好者(2016年6期)2016-12-21
中国铸造装备与技术(2015年5期)2015-12-10
