
不同碳源条件下里氏木霉的分泌蛋白比较分析
2018-11-07 06:46:30夏俊
生物化工 2018年5期
夏俊
(江西省科学院应用化学研究所,江西南昌330096)
纤维素是自然界中最丰富的生物质,是在生物产业中最具潜力的可再生和可持续能源[1-2]。木质纤维素是由纤维素、半纤维素、果胶和木质素组成的高聚物[3],其多糖成分可被酶降解为单糖和可发酵性糖[4]。然而,木质纤维素的应用和发展因植物细胞壁的坚固性而受到限制。de Souza等[5]人发现,甘蔗组织由约30%的纤维素、60%的半纤维素以及10%的果胶和木质素组成。对木质纤维素生物合成机制的研究表明,来自27个家族的356种糖基转移酶被预测参与细胞壁多糖糖苷键的形成[6]。因此,切断多糖糖苷键需要不同的糖苷水解酶。
至今已报道有多种真菌具有降解木质纤维素转化为可发酵糖的能力。里氏木霉(Trichoderma reesei)是一种可高效生产纤维素酶和半纤维素酶并协同降解木质纤维素的丝状真菌,它可产生大量纤维素酶以水解纤维素及其衍生物中的β-1,4糖苷键,产生的葡萄糖为真菌提供易于生长的碳源[7]。因对其生物应用的关注,里氏木霉中最丰富的纤维水解酶已成为结构和遗传研究的对象[8-9],包括两种纤维二糖水解酶(Cel7A和Cel6A,E.C. 3.2.1.91)、五种内切葡聚糖酶(Cel7B、Cel5A、Cel12A、Cel61A和Cel45A,EC 3.2.1.4)以及两种β-葡萄糖苷酶(BglI和BglII,EC 3.2.1.21)[10]。里氏木霉的半纤维素降解系统也由很多种酶组成,包括内切-1,4 -β-木聚糖酶(XYNI, 2, 3和4, EC 3.2.1.8)、甘露聚糖(MANI,EC 3.2.1.78)、乙酰聚木糖酯酶(AXEI,EC 3.1.1.72)、α-半乳糖苷酶(BGAI,EC 3.2.1.22)和阿拉伯呋喃糖苷酶(ABFI,EC 3.2.1.55)[11-12]。
研究表明,特异蛋白的mRNA表达水平与其实际表达水平没有明显的相关性[13]。因此,在蛋白质组学水平研究真菌的降解能力具有更大的现实意义。在过去的几十年,由于基因组数据的增加以及蛋白质组学技术的快速发展,蛋白质组学已成为组学研究的必要组成部分[14]。同时,蛋白质组学在丝状真菌降解木质纤维素的研究中也发挥了重要作用,进一步阐明了高效降解复杂底物所需的完整酶系。在转录水平上,里氏木霉的基因表达受到不同碳源的调控[15],但在分泌组水平上的相关研究还比较缺乏。本研究旨在分析和比较不同碳源条件下里氏木霉中分泌的纤维素降解蛋白,为合理、高效地转化由特定多糖组成的木质纤维素生物质提供依据。
1 材料与方法
1.1 培养基配方
无机盐溶液:NaNO36 g/L,KH2PO41.5 g/L,KCl 0.5 g/L,MgSO40.5 g/L,ZnSO4▪7H2O4.4 g/L,MnCl2▪4H2O 1.0 g/L,CoCl2▪6H2O0.32 g/L,CuSO4▪5H2O0.315 g/L,CaCl2▪2H2O1.47 g/L,FeSO4▪7H2O1 g/L。
马铃薯葡萄糖琼脂(PDA)培养基:马铃薯200 g/L,葡萄糖20 g/L,琼脂 15g/L。
玉米麦麸培养基(CSWB):玉米秸秆粉与麦麸以6∶4比例混合,每100g混合物粉末加200mL无机盐溶液用于固态发酵。
甘蔗渣培养基(SEB):甘蔗渣用压力为14kg/cm2蒸汽处理8min,用去离子水洗涤直至无还原糖残留,然后40℃干燥数天。每100g甘蔗渣加200mL无机盐溶液用于固态发酵。
甘蔗秸秆培养基(SC):将甘蔗秸秆粉碎成2mm左右的粉末,80%乙醇洗涤6遍再用去离子水洗除乙醇,干燥。每100g甘蔗秸秆加200mL无机盐溶液用于固态发酵。
1.2 固态发酵与样品收集
里氏木霉Rut-C30在PDA培养基中30℃培养,产孢后收集孢子悬浮于无菌水中,浓度约为7.0×107/mL。分别取200μL孢液接种于含30gCSWB培养基、SEB培养基和SC培养基的三角瓶中并于30℃培养5天后收集样品。向固体培养物中加入100mL无菌水并在4℃、200 r/min条件下萃取1h,随后将混合液10000r/min离心20min,保存上清液至4℃备用。
1.3 液相色谱-质谱(LC-MS)分析分泌组蛋白
培养5天后的胞外粗酶液通过超滤作用收集,再利用三氯乙酸沉淀,所得蛋白粉用双蒸水溶解。测定蛋白质浓度后,将10μL蛋白质含量小于100μg的蛋白质溶液加入到50μL含0.5 mol/LTris-HCl、2.75 mmol/L EDTA和6 mol/L盐酸胍的变性缓冲液中,然后将蛋白用1mol/L二硫苏糖醇还原处理2 h,再用1mol/L碘乙酰胺50μL黑暗中烷基化处理1 h。离心后加入360 μL 25 mmol/L的NH4HCO3,按1∶25(w/w)加入胰蛋白酶,于37℃下水浴中酶切过夜,用Sep-Pak C18柱脱盐后,将多肽样品溶解于含50%乙腈和0.1%三氟乙酸的水溶液中。
肽段的分析通过液相色谱-质谱联用完成。反相色谱柱采用MagicTMC18柱(75 μm ×10 cm,3 μm)。流动相A液为含2.0%(v/v)乙腈和0.1%(v/v)甲酸的水溶液,流动相B液为98%(v/v)乙腈和0.1%(v/v)甲酸的水溶液。2%~98%流动相B梯度洗脱流速0.3 μL/ min。质谱条件:喷雾电压2 kV,毛细管温度275 ℃,扫描范围400~1800 m/z,动态排除时间60 s,质量数据由软件Xcalibur 2.2.0采集处理。每个样品进行三次技术重复。
1.4 质谱数据的数据库搜索
采集的质谱数据用Proteome Discoverer software 1.4软件根据SEQUEST算法搜索里氏木霉的蛋白数据库(http://www.uniprot.org)。数据库搜索条件为:(1)胰蛋白酶漏切位点为2;(2)肽段碎片最大质量误差为0.8 Da;(3)蛋氨酸氧化作为动态修饰,半胱氨酸残基的氨基甲酰作为固定修饰物;(4)肽段匹配假阳性率为1%;(5)至少有6个碱基有95%以上确定性(q≤0.05)。
使用无标记的定量方法,通过肽谱匹配(PSMS)对蛋白质的相对丰度进行表征。所有蛋白质均使用SigalP 4.1预测分泌信号。
2 结果与分析
经筛选后,CSWB、SEB和SC三种培养基中的T. reesei分别含有分泌蛋白199、161、183个。如图1所示,分泌蛋白可分为纤维素酶、半纤维素酶、果胶酶、酯酶、淀粉酶、蛋白酶、几丁质酶、脂肪酶以及其他糖苷水解酶和一些未知功能的酶。T. reesei在三种培养基中的纤维素酶含量相近,CSWB和SC中的半纤维素酶含量比SEB高,但果胶酶含量都比SEB少。另外,半纤维素酶又可以分为木聚糖酶、1,3-1,4-β-葡聚糖酶、木葡聚糖酶和甘露聚糖酶等。T. reesei在三种培养基中的木葡聚糖酶含量相近,在CSWB和SC中分泌的木聚糖酶比SEB多,在CSWB中的1,3-1,4-β-葡聚糖酶比SEB和SC少。T. reesei在CSWB中分泌的甘露聚糖酶最多,其次是SC,而SEB中没有发现甘露聚糖酶。
2.1 与纤维素降解相关的酶
纤维素的完全降解需要外切葡聚糖酶(纤维二糖水解酶,CBHs)、内切葡聚糖酶(EGLs)、β-葡萄糖苷酶(BGLs)和其他辅助酶的协同作用。如表1所示,在CSWB、SEB和SC中均找到2种外切葡聚糖酶。在CSWB和SC中分别发现有3种和2种内切葡聚糖酶,而SEB中没有,但SEB中有4种β-葡萄糖苷酶,比CSWB和SC多。另外,多糖单加氧酶和膨胀素在CSWB中都发现有2种,在SEB和SC中都只有1种。
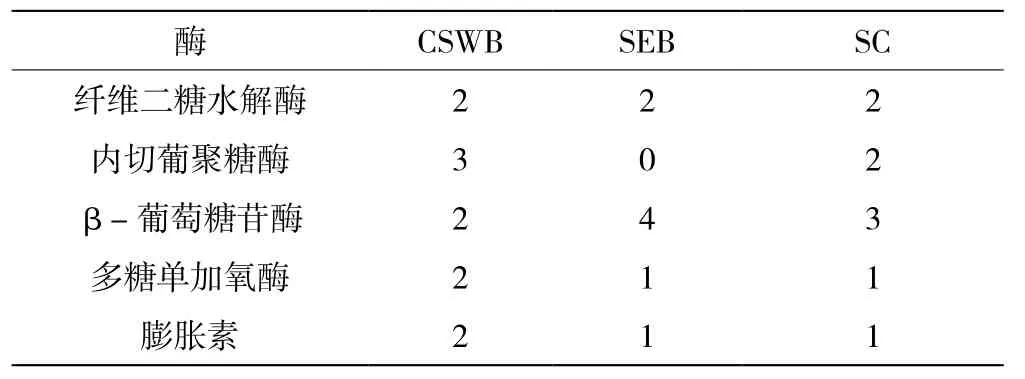
表1 各培养基中T. reesei分泌的纤维素降解酶数量
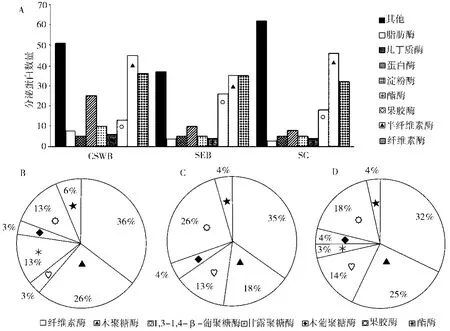
图1 不同培养基中T. reesei分泌组蛋白分类
2.2 与1,3-1,4-β-葡聚糖降解相关的酶
1,3-1,4-β-葡聚糖的完全降解需要破坏β-1,3和β-1,4糖苷键。本研究发现,T. reesei在SC中分泌较多的1,3-1,4-β-葡聚糖酶,包括1个1,4-β-D-内切葡聚糖酶、1个1,3-β-外切葡聚糖酶和2个1,3-β-葡糖苷酶。而在CSWB中只找到1个1,4-β-内切葡聚糖酶(表2)。
2.3 与木聚糖降解相关的酶
在CSWB和SC两种培养条件下的T. reesei分泌蛋白中都检测到了所有的主链酶(β-1,4-内切木聚糖酶和β-木糖苷酶)和支链酶(阿拉伯糖苷酶、葡糖醛酸酶和乙酰木聚糖酯酶),但在两者中的含量有所差异(表2)。CSWB和SC中的T. reesei均含有2个β-1,4-内切木聚糖酶和1个葡糖醛酸酶。另外,CSWB中T. reesei含1个β-木糖苷酶、2个乙酰木聚糖酯酶和2个阿拉伯糖苷酶,然与之相反,SC中的T. reesei却含有2个β-木糖苷酶、1个乙酰木聚糖酯酶和1个阿拉伯糖苷酶。在SEB培养基中,只发现2个β-1,4-内切木聚糖酶、1个β-木糖苷酶和1个乙酰木聚糖酯酶,而没有葡糖醛酸酶和阿拉伯糖苷酶。
2.4 与木葡聚糖降解相关的酶
木葡聚糖酶的完全降解依靠主链和支链酶的协同作用,包括一个降解主链的β-1,4-内切葡聚糖酶和11个支链酶(α-木糖苷酶、α-岩藻糖苷酶、β-1,4-牛乳糖苷酶和α-阿拉伯糖苷酶)。在CSWB、SEB和SC三种培养基中的T. reesei都含有β-1,4-内切葡聚糖酶,但除了在CSWB中含有1个β-1,4-牛乳糖苷酶外,三种培养基中都没有发现其他参与降解木葡聚糖的酶。
2.5 与甘露聚糖降解相关的酶
降解甘露聚糖的主链酶和支链酶,包括2个β-1,4-内切甘露糖酶、1个α-1,4-甘露糖苷酶和1个α-1,4-牛乳糖苷酶,都存在于CSWB培养基中;但SC中只有β-1,4-内切甘露糖酶而SEB没有任何参与甘露聚糖降解的酶(表2)。

表2 各培养基中T. reesei分泌的半纤维素降解相关酶的数量差异
3 讨论
本文分析和比较了在玉米麦麸培养基、甘蔗渣培养基和甘蔗秸秆培养基三种不同碳源条件下里氏木霉中分泌的纤维素降解蛋白。结果表明,提供不同碳源时,里氏木霉中的酶表达差异较大。麦麸中含有45%~50%的纤维素、半纤维素等成分;甘蔗秸秆中大约含有59%的全纤维素和20%的木质素;甘蔗渣的主要成分也是纤维素、半纤维素、木质素,其中半纤维素大约20.6%、木质素18.6%。纤维素在甘蔗渣里的含量大约为35.4%。因此,里氏木霉中的部分纤维素酶和半纤维素酶是诱导表达的,培养基中的不同碳源成分会诱导里氏木霉分泌所需的酶。
猜你喜欢
中国特种设备安全(2021年7期)2022-01-19 05:07:32
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:34:24
原子与分子物理学报(2021年1期)2021-03-29 07:28:26
食品与机械(2019年1期)2019-03-30 01:14:40
天然产物研究与开发(2018年8期)2018-09-10 05:48:24
天然产物研究与开发(2018年4期)2018-05-07 06:47:45
中国酿造(2014年9期)2014-03-11 20:21:06
中成药(2014年9期)2014-02-28 22:28:55
湖南农业科学(2014年5期)2014-02-27 14:29:42
