
抗双酰胺类杀虫剂小菜蛾的cDNA代表性差异分析
2018-11-05 04:02王莹莹李广阅徐巨龙孙诗晴薛超彬
山东农业科学 2018年10期
王莹莹,李广阅,2,徐巨龙,孙诗晴,薛超彬
(1.山东农业大学植物保护学院/农药毒理与应用技术省级重点实验室,山东 泰安 271018;2.山东鼎益生态农业有限公司,山东 临沂 276000)
近年来,随着氯虫苯甲酰胺和氟苯虫酰胺等双酰胺类杀虫剂的大量使用,小菜蛾对该类药剂的抗性问题也变得十分突出,尤其是在亚热带地区更为严重[1]。在菲律宾、泰国和中国的南方较早发现了高抗氯虫苯甲酰胺小菜蛾田间种群[2,3],与敏感品系/种群相比,采自泰国的田间抗性种群具有超过200倍的抗性[2];广东增城的抗性种群具有2 140倍的抗性[3];菲律宾的种群(Sudlon)具有超过4 100倍的抗性[2],近期的研究表明该Sudlon种群对氯虫苯甲酰胺的抗性倍数高达10 000[1];在巴西发现了对氯虫苯甲酰胺具有22 793倍的田间抗性种群[4]。
研究发现,泰国和菲律宾的小菜蛾田间高抗种群中鱼尼丁受体基因(PxRyR)的4 946位点均存在一个甘氨酸(G)到谷氨酸(E)的突变[2,5],被认为是导致小菜蛾对氯虫苯甲酰胺产生抗性的主要原因。本课题组在广东增城地区的田间高抗小菜蛾种群中也发现了该突变位点,位于受体基因羧基末端的跨膜区附近,且在所有昆虫鱼尼丁受体中高度保守[6]。同时,我们对不同抗性品系/种群小菜蛾进行了抗性频率检测,发现增城地区的抗性基因突变频率高达90%以上[7]。此外,解毒酶活性的提高,如细胞色素P450单加氧酶(P450)、羧酸酯酶(CarE)和谷胱甘肽S-转移酶(GSTs)活性的提高[8,9],PxRyR基因中4个位点的协同突变[10]以及PxRyR基因mRNA转录表达变化等[7,11,12],也是小菜蛾对氯虫苯甲酰胺产生抗性的重要机制,可见,小菜蛾对氯虫苯甲酰胺产生抗性的机制多样且较为复杂。
本试验在前期研究基础之上,利用小菜蛾敏感品系cDNA作为“驱动子”来源,小菜蛾抗氯虫苯甲酰胺品系、抗氟苯虫酰胺品系的cDNA分别作为“检测子”来源,采用改进的cDNA代表性差异分析法(cDNA RDA),筛选小菜蛾抗药性品系和敏感品系间的差异性表达基因,为锁定抗性靶标基因,进一步揭示小菜蛾的抗性机制奠定基础。
1 材料与方法
1.1 供试材料
小菜蛾敏感品系(S)于2006年采自山东农业大学南校区实验站园,在室内不接触任何药剂,用小白菜苗长期饲养且稳定繁殖;抗氯虫苯甲酰胺品系(Rf)和抗氟苯虫酰胺品系(Rh)是小菜蛾S品系分别在氯虫苯甲酰胺、氟苯虫酰胺药剂的持续选择压力下筛选获得的抗性品系,抗性倍数分别为684.54和677.25倍[13]。
1.2 RNA提取
分别选取抗氯虫苯甲酰胺品系(Rf)、抗氟苯虫酰胺品系(Rh)和敏感品系(S)小菜蛾的4龄幼虫,迅速用液氮冷冻,用RNAiso Plus试剂盒(宝生物大连工程有限公司)提取RNA,并利用1.5%琼脂糖凝胶电泳检测。采用PolyATtract®mRNA 纯化系统 Ⅲ (Promega,Madison,美国)进行小菜蛾mRNA的纯化。
1.3 小菜蛾双链cDNA的合成
采用RTAid Prem DS cDNA Synth Kit10 rxn (赛默飞世尔科技,北京)进行小菜蛾双链cDNA的合成。其中采用苯酚∶氯仿∶异戊醇(25∶24∶1)抽提双链cDNA,在30 μL 10 mol/L乙酸铵和250 μL乙醇中沉淀,然后-20℃过夜。4℃、14 000 r/min离心15 min,将得到的沉淀用70%乙醇洗涤2次,并风干。-20℃保存在ddH2O中备用。
1.4 检测子(Testers)和驱动子(Drivers)制备
本试验采用的代表性差异分析法(RDA)参考Hubank和Schatz[14]介绍的方法。其中接头和引物选用了3对接头(由上海生工生物工程有限公司合成),每对接头均为24个碱基及12个碱基的寡核苷酸序列,其中的24个碱基作相应的引物。“R”接头用于制备扩增子,“J”及“N”接头用于杂交扩增反应,所用引物如表1所示。
取获得的cDNA样品2 μg经DpnⅡ(New England Biolabs,Ipswich,MA,USA)消化酶切处理,将酶切消化产物连接R24/R12接头,16℃过夜。为了获得足够的下游所需扩增子,PCR反应采用22个扩增循环。扩增前先于72℃孵育3 min,随后加入Taq聚合酶,进行22个循环:95℃ 1 min,72℃ 3 min,72℃延伸10 min。经凝胶电泳证实每个扩增子的大小。用DpnⅡ消化去除扩增子的R24引物,作为下游反应的切割驱动子。在J12引物的作用下,将J24引物连接到切割驱动子中,即可作为第一轮杂交的检测子。

表1 cDNA代表性差异分析的接头和引物
1.5 消减杂交与RDA差异片段的分离
第一轮杂交将50 ng J24连接的检测子与4 μg驱动子混合,将混合物用苯酚+氯仿+异戊醇(25∶24∶1)抽提,然后置于30 μL 10 mol/L乙酸铵和250 μL乙醇中,-70℃沉淀1 h。4℃、14 000 r/min离心15 min,所得沉淀用70%乙醇洗涤2次,风干。将所得沉淀溶于5 μL杂交缓冲液(30 mmol/L EPPS pH 8.0,30 mmol/L EDTA pH 8.0,ρ=0.1 g/mL PEG8000)中, 37℃温育5 min,涡旋、离心。样品用35 μL矿物油覆盖,然后加热至95℃ 3 min进行变性,冷却至67℃,并将1 μL 1 mol/L的NaCl加入到DNA中,之后,样品在67℃中温育20 h。孵育后去除矿物油,用TE(Tris-EDTA)进行稀释,在下游PCR反应中使用5 μL杂交产物。杂交产物PCR扩增前,先72℃孵育3 min,加入Taq聚合酶。随后72℃ 5 min,加入引物(根据杂交轮次选用不同引物)。扩增条件:95℃ 1 min,然后70℃ 3 min(J24),共25个循环,最后72℃延伸10 min。为了将所得扩增产物用绿豆芽核酸酶(宝生物大连工程有限公司)消化,消除单链的扩增产物,本试验采用改进的PCR反应,即在7个循环后从PCR反应中取出10 μL试样,将其置于同样PCR反应条件中进行额外20个循环。按上述条件进行第二轮和第三轮的杂交和扩增,共进行三轮,至少两次重复,每轮反应中“驱动子”用量均为4 μg,检测子与驱动子的比例从第一轮的1∶100提高到第二轮的1∶800,直至第三轮的1∶20 000,其采用的接头引物分别为N24/N12和J24/J12。最终获得的基因片段在琼脂糖凝胶上呈现清晰条带,并进行测序分析。
2 结果与分析
2.1 cDNA RDA 差异片段的获得
小菜蛾S品系、Rf和Rh抗性品系的cDNA经DpnⅡ酶切后,在T4连接酶的作用下两端连上R接头,经PCR扩增,制备得到驱动子和检测子,琼脂糖凝胶电泳检测到200~750 bp的片段(图1)。分析三轮消减杂交片段,发现第一轮杂交产物背景较高并有严重的拖尾现象,说明除了选择性扩增之外仍有大量的相同基因未能被消减。随着消减杂交次数的增加,所获得的差异性片段逐渐减少,在第二轮杂交后出现明显的差异条带,第三轮消减杂交之后,主要差异条带得到了进一步的增强,大约在150~200 bp之间(图2)。
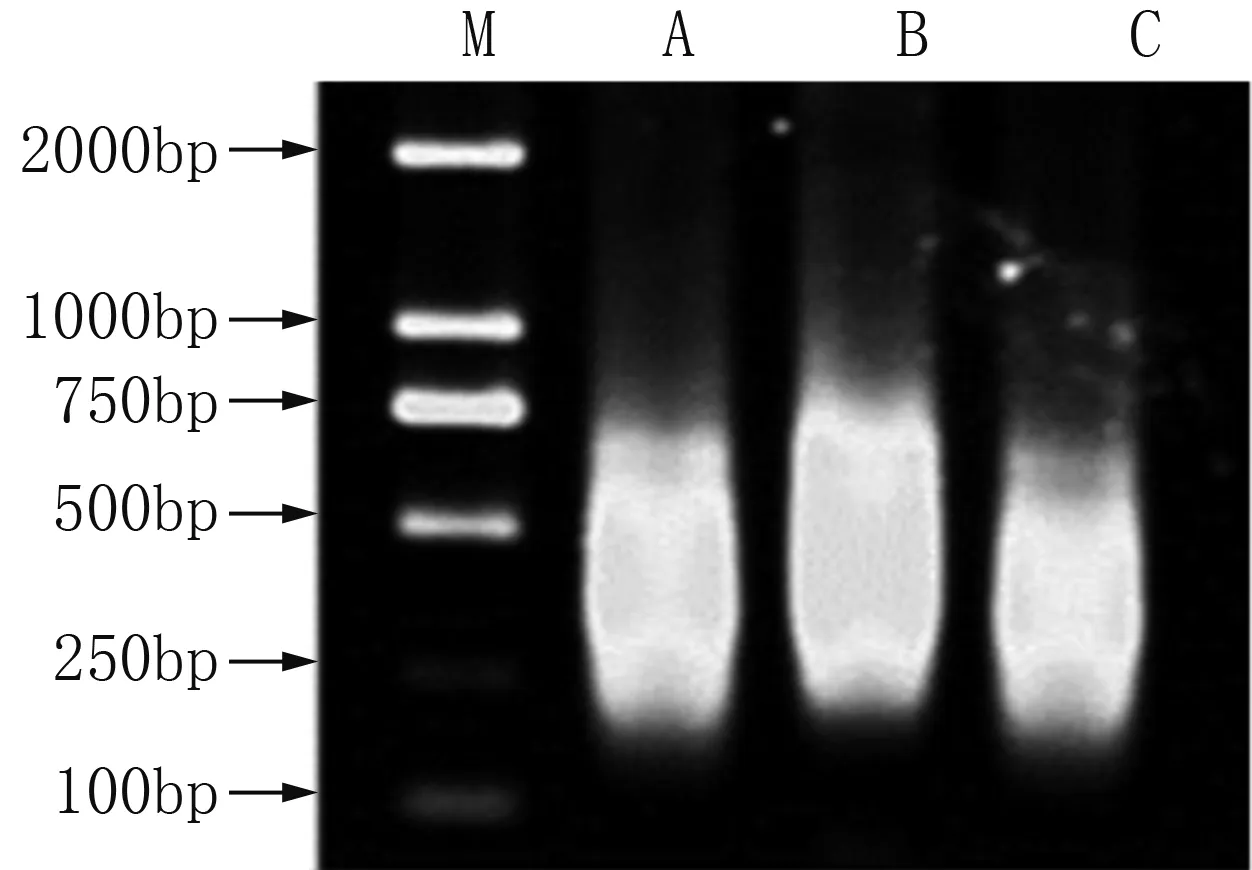
M为DNA Marker 2000;A为敏感品系;B为抗氯虫苯甲酰胺抗性品系;C为抗氟虫双酰胺抗性品系。
2.2 差异性片段的序列分析
将差异扩增片段克隆于pSIMPLE-18 vector(TaKaRa)载体上,转入菌株、氨苄筛选,菌落PCR鉴定阳性克隆,随机挑取12个菌落进行测序,将所得序列分别用NCBI BLAST 进行同源性比较。
小菜蛾抗氯虫苯甲酰胺品系获得9个cDNA RDA序列,其中有2条与28S rRNA基因有较高同源性,分别命名为DP3-1和DP3-2,同时发现有1个序列与已知任何序列无同源性,命名为DP3-3,可能为未知基因,其它序列均为重复序列。小菜蛾抗氟苯虫酰胺品系获得10个序列,其中有2个序列为DP3-1和DP3-2,同时也发现1个未知序列,但不同于DP3-3,命名为DP3-4(表2)。
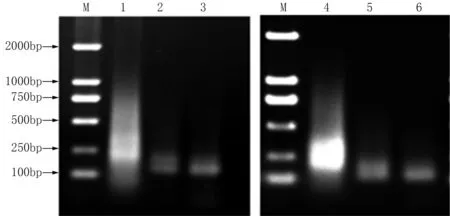
M为DNA Marker 2000;1~3为抗氯虫苯甲酰胺品系第一、二、三轮的杂交产物;4~6为抗氟苯虫酰胺品系第一、二、三轮的杂交产物。

表2 cDNA RDA 获得的差异性片段序列
2.3 差异性片段的同源性比较
DP3-1经NCBI BLAST同源性比较,其与家蚕28S rRNA的同源性为98%,与棉铃虫细胞色素P450类TBP蛋白基因同源性为97%(表3)。研究发现棉铃虫P450基因与昆虫的抗药性相关,主要表现为该基因的表达量上调,并表现出较高的P450活性。DP3-2经NCBI BLAST同源性比较,其与家蚕28S rRNA基因同源性为99%,与赤拟谷盗的TcasGA2_TC010626蛋白基因同源性为93%(表3),但该蛋白的功能目前尚不明确。

表3 cDNA RDA 获得的差异性片段的同源性比较
注:a、b分别代表序列在Rf和Rh抗性品系中RDA克隆的数目。
3 讨论
cDNA RDA方法被广泛应用于小菜蛾差异性基因片段的研究中,李宗英等利用代表性差异分析技术在抗杀虫双近等位基因中得到了2条差异片段(150~300 bp),初步确认为2个新序列[15];张晓飞等采用代表性差异分析方法,研究了敏感品系和抗杀虫双近等基因系,得到了2个差异片段[16];王淑珍用cDNA 代表性差异分析技术从小菜蛾杀虫双抗性品系和敏感品系中得到了一个差异性片段(100~200 bp),该基因片段与28S rRNA基因有较高的同源性[17];刘白朵等通过双向cDNA RDA方法,从溴氰菊酯抗性品系小菜蛾和敏感品系小菜蛾中发现了一条序列与编码S3a蛋白的基因有较高的同源性[18]。由此可见,cDNA RDA方法在小菜蛾抗药性基因的筛选中发挥了重要的作用。
在消减杂交试验中,抑制性消减杂交(SSH) 和cDNA代表性差异性分析(cDNA RDA)是目前应用最广泛的方法,但是SSH更适合构建差异基因文库,因为该方法仅两轮杂交,当两个样本间的差异较小时假阳性率高,SSH中两次驱动子 cDNA使用量均过量,可能会掩盖检测子cDNA中某些有表达差别的基因,因此,有些差异表达的基因可能检测不到。本试验中两个样本之间遗传背景相同,差异性较小,所以更适合用cDNA 代表性差异分析方法。
本研究利用cDNA 代表性差异分析技术从抗氯虫苯甲酰胺和抗氟苯虫酰胺抗性品系中筛选出了2个差异基因片段DP3-1和DP3-2,同时得到了2个未知序列,DP3-1与棉铃虫的细胞色素P450类TBP蛋白基因有97%的同源性。在已报道消减杂交的试验中都得到了细胞色素P450类TBP蛋白,何鹏通过抑制性消减杂交方法在低盐胁迫下的橡胶树中得到了细胞色素P450类TBP蛋白基因[19];张建清通过抑制性消减杂交的方法从球孢白僵菌侵染昆虫和附着胞形成时期受BbMPK1 MAPK调控的下游基因分离得到了烟草细胞色素蛋白(cytochrome P450 like TBP)[20];李玲等在桃花芽休眠解除SSH差减文库中得到了红花烟草细胞色素 P450类TBP[21]。研究表明,昆虫的细胞色素P450与杀虫剂的抗药性有关,细胞色素P450酶系是与代谢抗性相关的主要解毒酶系之一,其抗药性原理主要是由解毒作用的增强而导致的,表现为相应的P450酶活性提高或其表达量的增强。本研究为后续深入研究小菜蛾对双酰胺类杀虫剂的抗性机制提供了新线索。
猜你喜欢
植物保护(2022年1期)2022-02-10
农业技术与装备(2022年11期)2022-01-01
右江民族医学院学报(2021年4期)2021-09-18
现代农药(2021年2期)2021-05-07
上海农业科技(2020年5期)2020-10-24
当代水产(2020年3期)2020-06-15
世界农药(2019年3期)2019-09-10
世界农药(2019年3期)2019-09-10
农药科学与管理(2019年10期)2019-04-20
蔬菜(2018年9期)2018-09-21
