
白念珠菌SIM1基因敲除与功能研究
2018-11-01 02:16张晓龙鲁仁义阎澜姜远英
中国真菌学杂志 2018年4期
张晓龙 鲁仁义 阎澜 姜远英
(中国人民解放军海军军医大学药学院新药研究中心,上海 200433)
白念珠菌是人体常见的条件致病型真菌,可呈现出酵母态、假菌丝态及菌丝态三种完全不同的状态[1]。在人体免疫力低下时,白念珠菌能够穿过人体上皮屏障、侵入深部黏膜并播散进入血液循环中,导致严重的系统性真菌感染[2]。目前,临床上白念珠菌感染面临着病死率高、耐药性强、特效药物少等棘手问题[3],因此,研究白念珠菌致病机理,对于开发新型抗真菌药物具有重要意义。
SUN基因家族是一组真菌特异性基因家族,其蛋白质在C末端结构域具有高度相似性[4]。在酿酒酵母中,Sun蛋白家族最初在鉴定出芽酵母酿酒酵母中的四个旁系同源基因时被识别,即SIM1,UTH1,NCA3和SUN4基因[5],这个家族的基因参与DNA复制、老化、线粒体合成和胞质分裂等多种细胞过程[6]。在白念珠菌中,SUN家族包括SUN41和SIM1两个基因。研究表明,sun41蛋白具有糖苷酶功能,参与白念珠菌形态、细胞壁以及生物被膜形成。SUN41基因缺失菌会出现细胞分离障碍现象,其菌丝及生物被膜形成产生缺陷,黏附宿主的能力也显著下降[7]。在本研究中,我们通过基因敲除获得了SUN基因家族另一基因SIM1的基因缺失菌株,并对其表型和功能进行了初步探究。
1 材料和方法
1.1 材料
菌株 菌株SN152 (亲本菌为SC5314,基因型为arg4Δ/arg4Δleu2Δ/leu2Δhis1Δ/his1ΔURA3/ura3Δ::imm434IRO1/iro1Δ::imm434)由美国加利福尼亚大学SuzanneM.Nobel教授馈赠。
试剂YeaStarGenomicDNAKit,购于ZymoResearch公司。YeastTransformationSystem2购于Clontech公司。DNAMarkerDL2000、DNAMarkerDL10000购于Takara公司。Goldview染料购于北京赛百盛公司。磷酸盐缓冲液 (phosphatebufferedsaline,PBS)购自上海博光公司。
药物 氟康唑、咪康唑、酮康唑、卡泊芬净、两性霉素B、特比萘芬、刚果红、荧光白购于Sigma公司,氯化锂购于中国医药集团上海化学试剂公司。
培养基YPD液体培养基 (1%酵母浸膏、2%蛋白胨、2%葡萄糖)、SDA固体培养基 (1%蛋白胨、4%葡萄糖、1.8%琼脂、各药物)、Spider液体培养基 (1%酵母浸膏、0.2%KH2PO4、1%甘露醇)、RPMI1640液体培养基 (1%RPMI1640、3.45%吗啉基丙磺酸、0.2%NaHCO3)、SC固体培养基 (2%葡萄糖、0.67%酵母氮源、1.8%琼脂、所需氨基酸)。
1.2 方法
菌株培养 于-80℃冰箱中将20%甘油冻存的白念珠菌SN152菌液接种至SDA固体培养基平板中,30℃倒置培养48h后挑取单个菌落继续接种在SDA平板上,30℃再次活化48h。挑取单克隆接种于1mLYPD液体培养基,在30℃、200r/min振荡培养16h后用于实验。
SIM1基因敲除实验 使用醋酸锂化学转染[8]的方法,利用Overlap-PCR介导的HIS1-LEU2基因策略[9-10],将C.dubliniensis中的HIS1和C.maltosa中的LEU2两个标记同源重组替换SN152中SIM1基因,构建双臂敲除菌,构建示意图见图1。首先以SN152基因组DNA为模板,SIM1_up_F、SIM1_up_R和SIM1_down_F、SIM1_down_R分别为引物PCR扩增SIM1基因上、下游片段。再用P2、P5分别以pSN52、pSN40为模板PCR扩增C.d HIS1marker和C.m LEU2marker。以SIM1基因上游、下游、HIS1或LEU2marker3个片段为模板,利用SIM1_up_F、SIM1_down_R为引物融合PCR扩增得到含有HIS1或LEU2的融合重组片段。然后利用醋酸锂法将HIS1融合片段转染进入白念珠菌SN152中,涂布于选择性培养基SC-His平板上,48h后挑取菌落,抽提基因组DNA,PCR鉴定得到SIM1的单臂缺失菌,同样方法转染LEU2融合片段至SIM1单臂缺失菌中,在SC-His-Leu培养基上筛选生长,经PCR鉴定得到SIM1基因缺失突变菌SIM1Δ/Δ。菌株构建和鉴定所需引物应用NCBIPrimerdesigningtool设计,由上海生工公司合成,-20℃保存,各引物序列见表1。PCR产物经琼脂糖凝胶电泳鉴定。
生长曲线实验 从SDA平板挑取不同白念珠菌克隆至1mLYPD液体培养基,30℃、200r/min振荡培养16h。离心收集菌体,用无菌PBS溶液洗涤3次,并用新鲜YPD调整菌浓度为1×106cells/mL,测菌液OD630nm值记为0h。将菌液继续30℃、200r/min振荡培养,于不同时间点测量菌液OD630nm值。
菌丝诱导实验 活化菌株,用无菌PBS溶液洗涤3次,分别用RPMI1640、YPD+10%FBS、Spider液体培养基调整菌浓度为1.0×106cells/mL。将调整好的菌液分别加至12孔板并于37℃静置培养3h,通过显微镜观察菌丝形成情况。
黏附上皮细胞实验 活化菌株,用无菌PBS溶液洗涤3次,用MEM培养基 (含20%胎牛血清)重悬菌株,调整浓度为1×105cells/mL,然后稀释至终浓度为200cells/mL。待结肠癌上皮细胞 (Caco-2细胞)和口腔表皮样癌细胞 (KB细胞)在6孔板中融合生长至80%~90%时,加入1mL上述菌液,然后置于37℃、5%CO2细胞培养箱孵育1h,吸弃上清液,用无菌PBS溶液轻轻洗3次,以未用PBS漂洗的样本孔做对照。每孔加入1mL尚未凝固的YPD固体培养基 (45℃),于30℃倒置培养48h,记录生长的白念珠菌克隆数并计算黏附率。
生物被膜活性实验 活化菌株,用无菌PBS溶液洗涤3次,用RPMI1640培养基重悬菌株,调整浓度为1.0×106cells/mL。在96孔板中分别加入100μL调好浓度的菌液,于37℃培养箱静置培养24h,然后轻轻吸弃上清的RPMI1640培养基,用无菌PBS溶液轻轻洗涤3次,于每孔分别加入150μLXTT/menadione混合液,37℃避光静置孵育3h,最后吸取70μLXTT/menadione液体上清测OD495nm值。
Spotassay实验 活化菌株,用无菌PBS溶液洗涤3次,并调整菌液浓度至1×107cells/mL,然后用无菌PBS连续10倍倍比稀释5个浓度梯度。取3μL上述稀释菌液点到各含药SDA固体培养基上,将平板晾干,于30℃培养箱倒置培养48h,观察菌落状态并拍照。

表1 实验所用引物及序列
2 结 果
2.1 SIM1基因缺失菌构建成功
鉴定结果如图2所示。1%琼脂糖凝胶电泳成功验证出同源重组SIM1基因双臂敲除菌HIS1和LEU2标记的上下游片段:HIS1上游(HIS1 Left+ up-SIM1)、下游(HIS1 Right+ down-SIM1)、LEU2上游(LEU2 Left+ up-SIM1)、下游(LEU2 Right+ down-SIM1)四个片段,大小分别为1 022 bp、1 136 bp、654 bp和1 124 bp。而SIM1基因中两个ORF片段 (1_F+1_R,724bp)、(2_F+2_R,769bp)均没有出现,说明白念珠菌SIM1Δ/Δ构建成功。
2.2 SIM1基因缺失后不影响白念珠菌的生长、繁殖
在宿主体内侵袭性生长、繁殖是白念珠菌逃逸宿主免疫攻击,难以被宿主清除的主要方式之一。通过测定生长曲线可以考察SIM1基因缺失后对白念珠菌生长、繁殖的影响,结果如图3所示,SIM1基因缺失菌SIM1Δ/Δ与亲本菌SN152相比,生长繁殖速度无明显变化。说明SIM1基因缺失后白念珠菌的生长、繁殖不受影响。
2.3 SIM1基因缺失后不影响白念珠菌菌丝形成
一般情况下,白念珠菌以酵母态与宿主共生,当宿主免疫功能降低时,白念珠菌转化为具有侵袭性的菌丝态生长,对宿主造成侵袭性感染[11]。本实验通过在不同液体培养基中诱导白念珠菌形成菌丝来考察SIM1基因缺失后对白念珠菌菌丝形成能力的影响。结果如图4显示,与亲本菌SN152相比,SIM1基因缺失菌SIM1Δ/Δ的菌丝形成情况无明显差异。
2.4 SIM1基因缺失导致白念珠菌对宿主上皮细胞的黏附能力降低
黏附是白念珠菌感染宿主过程中的第1步,也是影响白念珠菌致病力的重要因素。本实验检测了亲本菌SN152和SIM1基因缺失菌SIM1Δ/Δ对宿主上皮细胞的黏附能力。结果如图5所示,亲本菌SN152和SIM1基因缺失菌SIM1Δ/Δ对Caco-2细胞的黏附率分别为76.50%和56.63%;对KB细胞的黏附率分别为78.87%和56.33%。与亲本菌相比,白念珠菌SIM1Δ/Δ对上皮细胞的黏附能力显著降低 (P<0.05)。
2.5 SIM1基因缺失后不影响白念珠菌被膜形成
生物被膜与白念珠菌感染密切相关,能大大增强白念珠菌对抗真菌药物的耐药性和对宿主免疫防御的抵御能力[12]。本实验通过XTT法[13]考察亲本菌SN152和SIM1基因缺失菌SIM1Δ/Δ在96孔板内壁上形成生物被膜的活性。结果如图6所示,以亲本菌SN152形成的生物被膜为参照,SIM1Δ/Δ菌的生物被膜与其没有显著差异。

图1白念珠菌SIM1基因敲除菌株构建示意图图2PCR验证SIM1基因双臂敲除菌图3SIM1基因缺失后不影响白念珠菌生长繁殖图4SIM1基因缺失后不影响白念珠菌菌丝形成图5SIM1基因缺失后白念珠菌对宿主上皮细胞黏附能力降低。实验重复3次,数据采用mean±SD表示,t检验分析两组间黏附率均数的统计学差异 (*.P<0.05)图6SIM1基因缺失后不影响白念珠菌被膜形成。实验重复3次,数据采用mean±SD表示,t检验分析两组菌被膜形成率差异的统计学意义
Fig.1Schematic diagram of constructing deletion mutants forCandidaalbicansSIM1Fig.2PCR confirming disruption ofSIM1 two allelesFig.3Deletion mutants ofSIM1 did not affect the growth and reproductionFig.4Deletion mutants ofSIM1 did not affect the hyphae formationFig.5Deletion mutants ofSIM1 impair the adhesive ability to Caco-2 cells and KB cellsFig.6Deletion mutants ofSIM1 did not affect the biofilms formation
2.6 SIM1基因缺失后白念珠菌对刚果红、荧光白的药物敏感性增加
细胞壁是白念珠菌维持细胞形态、平衡渗透压,保护自身免于环境胁迫的重要细胞器。目前,临床上常见的抗真菌药的作用机制主要通过破坏细胞壁及细胞膜的结构、通透性,达到杀灭或抑制真菌的目的[14]。本实验通过spot assay实验来考察SIM1基因缺失后白念珠菌药物敏感性的变化。结果如图7所示,与亲本菌相比,SIM1基因缺失菌在刚果红 (100 μg/mL)和荧光素白 (50 μg/mL)的含药培养基上生长受到明显抑制。刚果红和荧光素白的作用靶点集中在细胞壁上,说明SIM1基因缺失后导致白念珠菌细胞壁发生改变,对药物的敏感性增加。
3 讨 论
基因敲除技术是研究白念珠菌基因功能的常用方法。基因敲除技术从最早的只有以URA3为筛选标记的Ura-Blast策略[15],发展到以HIS、LEU2和ARG4为筛选标记的HIS1-LEU2-ARG4基因敲除策略[9]、以诺尔斯菌素为筛选标记 (SAT1)的诺尔斯菌素基因敲除法[16]等多种实验方法。本实验所采用的HIS1-LEU2策略不仅回避了URA3对白念珠菌毒力的影响,且具有筛选步骤少、转染效率高等特点。
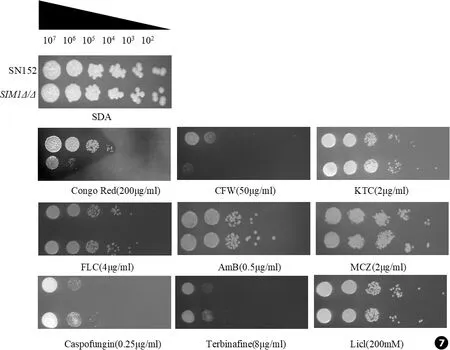
图7 SIM1基因缺失后影响白念珠菌对部分外界应激的敏感性
Fig.7Deletion mutants ofSIM1 affect the sensitivity to some external stress (CFW.Calcofluor white,KTC.Ketoconazole,FLC.Fluconazole,AmB.Amphotericin B,MCZ.Miconazole)
有研究表明,白念珠菌SUN基因家族中SUN41基因功能涉及母细胞和子细胞分离时广泛的细胞壁重塑,SUN41基因失活会导致白念珠菌细胞分离障碍和菌丝形成缺陷[7]。本实验结果表明SIM1基因失活仅造成白念珠菌黏附能力下降和对刚果红、荧光素白的的敏感性增加,而不会对白念珠菌细胞分离障碍和菌丝形成造成影响。在酿酒酵母和裂殖酵母中,SUN基因已证实与RAM-Ace2p信号通路相关联,该信号通路通过调节子细胞中编码细胞壁降解酶基因的特异性表达促进母细胞细胞壁降解,在细胞分离中发挥着重要作用[17]。酿酒酵母中SIM1基因通过改变细胞壁成分维持细胞分离后母细胞细胞壁的完整性,不影响子细胞Ace2p的特异性定位也不抑制细胞分离[18]。白念珠菌的SIM1基因与酿酒酵母的SIM1基因同源,具有58.8%相似性。在本实验中,SIM1基因缺失后白念珠菌的生长、菌丝形成、细胞分离均不受影响,而对刚果红、荧光白的药物敏感性增加。种种迹象表明白念珠菌SIM1基因功能与酿酒酵母SIM1基因功能极为相似。值得一提的是,刚果红和荧光白两种药物均与几丁质的生物合成相关。白念珠菌SIM1基因是否通过调节几丁质生物合成从而改变细胞壁成分,并进一步影响白念珠菌对宿主的黏附值得进一步研究。
综上所述,本研究成功构建了白念珠菌SUN基因家族另一基因SIM1的基因缺失菌,并对其功能进行了初步探究,为进一步阐释SUN基因家族在白念珠菌生长、传代和与宿主相互作用中的功能提供了基础。
猜你喜欢
首都食品与医药(2022年11期)2022-06-06
中国土壤与肥料(2021年5期)2021-12-02
科学(2020年3期)2020-11-26
疯狂英语·新悦读(2020年7期)2020-07-30
当代水产(2020年3期)2020-06-15
医药前沿(2020年4期)2020-04-27
癌变·畸变·突变(2020年1期)2020-02-12
中国卫生标准管理(2015年24期)2016-01-14
小星星·阅读100分(高年级)(2015年11期)2015-11-28
医学研究杂志(2015年12期)2015-06-10
