
棉蚜腺苷三磷酸酶E亚基基因RNAi载体的构建及其抗蚜性分析
2018-11-01 09:29罗晓丽张安红肖娟丽王志安陈晓英吴家和
棉花学报 2018年5期
罗晓丽 ,张安红 ,肖娟丽 ,王志安 ,陈晓英 ,吴家和 *
(1.山西农业科学院棉花研究所/棉花种质资源利用与分子设计育种山西省重点实验室,山西 运城044000;2.中国科学院微生物研究所/植物基因组国家重点实验室,北京100101)
棉蚜(Aphis gossypiiGlover)俗称腻虫,是同翅目蚜科的1种昆虫,是棉花苗期的主要害虫之一。棉蚜为世界性棉花害虫,在中国所有棉区都有分布,西北地区尤为严重。棉蚜寄主植物广泛,包括石榴、花椒、木槿、鼠李属、棉花、瓜类等。棉蚜是刺吸式昆虫,吸食棉花的汁液,致使其产量下降、品质降低[1]。此外,它还是传播植物病毒的优良媒介。使用农药虽然可以在短时间内有效地防治,但是极易使蚜虫产生抗药性。同时,农药会杀死蚜虫的天敌,这些天敌种群恢复明显滞后于蚜虫,导致蚜虫种群的再次爆发。此外,农药的不合理使用会污染环境,与当前环境保护、绿色生态农业的主流相悖[2]。因此,培育抗蚜棉花品种显得十分迫切。虽然育种工作者在形态育种上取得了一定的避虫进展,但是并没有选育出真正意义上的抗蚜虫品种[3]。当前转基因抗虫棉在抗棉铃虫、红铃虫方面产生了巨大效益,然而对刺吸性害虫抗性较差或根本没有抗性[2,4]。因此,寻求新的技术手段来创造抗蚜棉花材料成为棉花抗蚜育种亟待解决的新课题。
近十几年来,人们一直在进行抗蚜基因挖掘及棉花抗蚜研究。来自番茄的Mi-1.2基因对马铃薯蚜虫(Myzus persicaeSulzer)有一定的抗性[5];从甜瓜中克隆的Vat基因对棉蚜具有较好的抗性[6];从大豆中分离的 Rag 基因(Rag1、Rag2、Rag3)对大豆蚜虫(Aphis glycinesMatsumura)具有很好地抗性,孟山都公司已将Rag1基因在大豆中过量表达,获得了抗蚜性增强的大豆。Rag3具有更广泛的蚜虫抗性[7]。另外,从苹果中分离的抗苹果红西圆尾蚜(Dysaphis devectaWalker)基因Sd-1和从野生莴苣中分离的对莴苣蚜虫(Nasonovia ribisnigr)具有专一抗性的Nr抗蚜基因[8]。目前通过基因工程防治蚜虫虽然有了较大发展,获得了一些抗蚜虫基因,但是这些抗蚜基因对蚜虫仅有抑制作用,不能满足生产实践中对蚜虫防治的要求。对一些广谱抗蚜基因的研究报道也较多,如细胞分裂素基因,过量表达该基因影响蚜虫的取食[9],植物凝集素基因也是1类广谱抗蚜基因,如来自雪花莲凝集素基因(Galanthus nivalis Agglutinin,GNA)在烟草中表达能够增强对桃蚜的抗性[10],转大蒜甘露糖特异性凝集素基因的水稻对刺吸类昆虫的抗性增强[11]。然而,目前还没有见到转上述基因植物大田推广应用的报道。
RNA沉默是双链 RNA (Double stranded RNA,dsRNA)被特异的核酸酶降解,产生小干扰RNA (Small interfering RNA,siRNA), 这些 siRNA与同源的靶RNA互补结合,特异降解靶RNA,从而抑制基因表达。最近研究发现干扰性RNA(dsRNA、siRNA)可以通过抑制线虫、鞘翅目昆虫消化道基因的表达[12]。液泡型腺苷三磷酸酶(Vacuolar ATPase,V-ATPase)存在于所有真核细胞中,由多亚基装配,其中 E亚基(V-ATPase E)是1个具有丰富带电残基的亲水性蛋白,对酶与阳离子的结合起调节作用。在逆境胁迫响应过程中能保持生物体内能量代谢平衡,对生物体生长发育及存活至关重要。近几年随着ATPase亚基序列的公布,对各个亚基功能和抗逆性方面的研究也越来越多,在昆虫中的研究发现,敲除或敲低V-ATPase的各亚基基因常导致昆虫死亡。如通过 RNA 干扰(RNA interfering,RNAi)降低黑腹果蝇 (Drosophilamelanogaster), 赤拟谷盗(Tribolium castaneum),豌豆蚜(Acyrthosiphon pisum)和烟草天蛾(Manducasecta)的 V-ATPaseE 亚基[13],玉米根莹叶甲(Diabrotica virgifera)的 V-ATPaseA、D、E亚基的表达[14],都会导致死亡或对生长和/或生殖的产生其它负面影响。但是利用RNAi技术敲除棉蚜腺苷三磷酸酶E亚基基因(AgATPase E,本文简称AgApe)进行抗蚜虫方面的研究还少见报道。
本研究通过构建棉蚜腺苷三磷酸酶E亚基基因的RNAi载体并转化棉花对45个独立转化植株进行聚合酶链式反应(Polymerase chain reaction,PCR)筛查,获得 22个 PCR阳性株,进行插入拷贝数鉴定和表达量鉴定,选出4个转基因株系进行2年的网棚和大田抗蚜性鉴定。结果表明4个转基因株系对蚜虫具有显著的抗性。本研究建立了1种新的利用RNAi技术干扰AgATPase E基因代谢途径的抗蚜策略,也为棉花抗蚜育种提供了种质资源。
1 材料和方法
1.1 试验材料
试验所用转基因受体为的中棉所35,种子保存于山西农业科学院棉花研究所。采用肖松华等[15]的方法培养蚜虫,在隔离的温室种植常规棉花不做任何药剂处理,温室温度控制在20~27℃,通过定期浇水和温室内洒水将湿度控制在适宜的范围内。利用自然虫源接种棉蚜,让其自然繁殖,供网棚蚜虫抗性鉴定试验用虫。
1.2 RNAi表达载体构建
根据GenBank中的豌豆蚜虫基因组序列中腺苷三磷酸酶E亚基基因(基因登录号:CN582351),设计同源序列引物从棉蚜cDNA序列文库中扩增靶基因序列,将基因片段克隆到pGEM-T载体上并进行测序验证。以中国科学院微生物研究所植物基因组国家重点实验室的pIPK质粒为基础构建植物hpRNA表达载体[16],该质粒含有潮霉素抗性基因。用于靶基因的正义序列分别用EcoRⅠ和KpnⅠ插入到pIPK质粒上,然后利用BamHⅠ和XbaⅠ双酶切插入靶基因的反义片段,构建成植物表达载体pCAMBIA-HAN-dsAgA(图1)。表达载体转化到农杆菌菌株 EHA 105并测序验证[16]。
1.3 转基因再生植株培育和PCR分析
转化受体为中棉所35,具体技术和方法参照已报道的文献[17-18]。转基因棉花再生植株移栽到温室后,叶片涂抹潮霉素检测抗性,并提取DNA进行PCR分析。PCR阳性植株进行自交并收获种子。
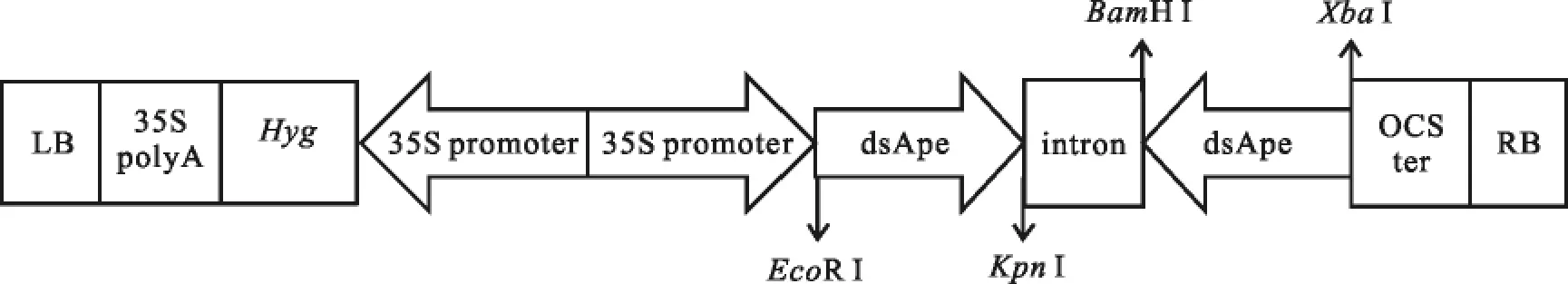
图1 植物表达载体pCAMBIA-HAN-dsAgA的结构示意图Fig.1 Schematic diagram of plant expression vector pCAMBIA-HAN-dsAgA
选取转基因再生株及其后代幼嫩叶片进行DNA提取。DNA的提取使用Promega公司植物DNA提取试剂盒 (离心柱法),提取DNA后用Nanodrop分光光度计(2000/2000C)进行浓度检测, 稀释成 100 μg·L-1作为 PCR的 DNA 模板。PCR 反应体系:10 μL PCR mix、0.5 μL DNA 模板、1.0 μL引物和 ddH2O 混合成 20 μL。 PCR 扩增程序为:95℃预变性 5 min,94℃变性 40 s,55℃退火 40 s,72℃延伸 30 s,35 循环,72℃延伸 5 min。引物序列见表1。阳性植株进行进一步的拷贝数和转录表达的检测。收获T0种子,在网室里进行后代的选育和分子鉴定。
1.4 Southern blot分析
对PCR阳性植株进行分子杂交试验,分析AgApe基因片段的拷贝数。用限制性内切酶HindIII酶切转基因棉花叶片总DNA,具体分子杂交方法参考相关文献[19],探针为上述PCR扩增的AgApe基因片段。
1.5 转录表达检测分析
利用反转录聚合酶链式反应 (Reverse transcription PCR,RT-PCR)方法对转基因植株及其后代转录表达进行分析。摘取转基因植株棉花幼嫩叶片,利用Promega公司RNA提取试剂盒进行总RNA的提取,采用Nanodrop测定RNA浓度。Promega公司SuperScriptⅢFirst-Strand Synthesis System试剂盒将 2 μg的 RNA逆转录成cDNA,用表1中的A-EcoRI-F和 A-KpnI-R引物进行PCR扩增。转化受体为阴性对照。棉花Actin基因作为内参,引物序列如表1所示。

表1 特异引物序列Table1 Sequence of specific primers
1.6 转基因植株抗蚜性分析
采用肖松华等[20-21]方法对转AgApe基因RNAi载体的棉花纯合系的抗蚜性进行分析。2015―2016年分别在网棚接虫鉴定和大田自然鉴定。网棚抗蚜性鉴定:纯合的转基因后代种植于网室,出苗后罩上网棚,网棚内无棉蚜天敌存在。3叶期(4月25日左右)实施接蚜处理,每株接材料1.1中培养的混合蚜(4~5头成蚜,其它为1~5日龄的若蚜)40头。于5叶期(5月10日左右)调查棉花蚜害级别,以受体品种为对照,计算蚜害指数及其比对照减退率(X)并进行评价,抗性鉴定分级评价标准如表2所示。

表2 转基因抗蚜棉对棉蚜抗性的分级标准Table2 Transgenic cotton aphid resistance to aphid grading standards
蚜害指数=∑(各级蚜害株数×相应级别)×100/(调查株数× 4)
蚜害指数减退率=(受体品种蚜害指数-鉴定材料蚜害指数)/受体品种蚜害指数×100%
大田自然鉴定蚜虫抗性:2015―2016连续2年转基因棉花种植于大田,苗期 (4月20日―5月20日)连续1个月不喷施农药,田间自然发生蚜虫危害,具体检测指标及评价方法同网棚。
2 结果分析
2.1 RNAi表达载体的构建和转基因棉花的获得
利用同源克隆的方法获得棉蚜的AgApe基因靶标序列,片段大小为282 bp的特异序列。对1.2构建好的载体质粒进行酶切验证(图2)和测序,能够检测到靶基因序列(ATGGAGTATTTTG AGCGCAAAGAGAAACAAGTGGAGTTACAGG CTTCGGTTCAATCATCTAATATGTTAAATCA AGCACGTCTTAAGACATTAAAAGTGCGAGA AGATCATGTCAGTGATGTATTAGATGAAGCA AGGAAGCGTCTGGTCAAAGTGACAAATAAC CCTGATTTGTACAGAGAAGTTCTTAGAAAAT TAATATTGCAAGCAATCTTACAGTTGTTGGA GAAAAATGTTACATTACGTGTTCGTGAAGTT GATTTATCAGTTGTGGAAGAATTA),序列一致,说明RNAi表达载体构建成功(图1)。
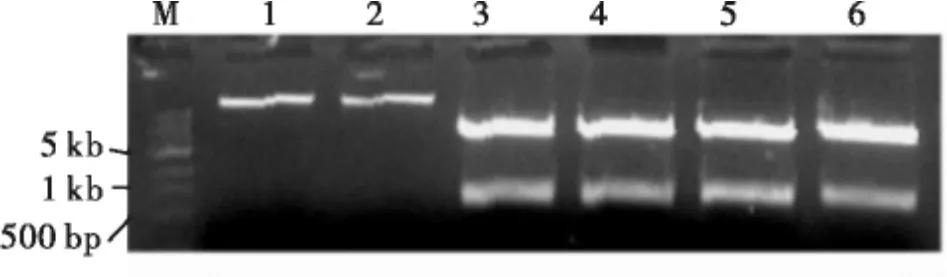
图2 植物表达载体pCAMBIA-HAN-dsAgA的酶切鉴定Fig.2 The digestion analysis of plant expressing vector pCAMBIA-HAN-dsAgA
图3是以中棉所35为转化受体,获得转基因植株的过程。共获得45株T0转基因再生植株,并移栽于温室进行正常管理。对这些转基因植株叶片中部涂抹 20 mg·L-1潮霉素,6~7 d后观察。结果显示:41个再生植株叶片涂抹部位呈现绿色,为潮霉素抗性植株;有 4株叶片涂抹部位呈现枯黄色为潮霉素阴性植株(图4)。
对45株转基因棉花材料进行PCR检测,结果发现有41株PCR呈阳性,与潮霉素涂抹实验结果一致。如图5所示,转基因植株能够扩增出950 bp左右条带,与质粒正对照扩增的条带大小一致,而转化受体材料没有扩增出相应的条带。对PCR产物进行测序分析,结果显示扩增的片段为AgApe基因片段(表1)。结果表明93.3%转基因植株为PCR阳性植株。共收获22株能够正常开花授粉的转基因阳性植株种子。
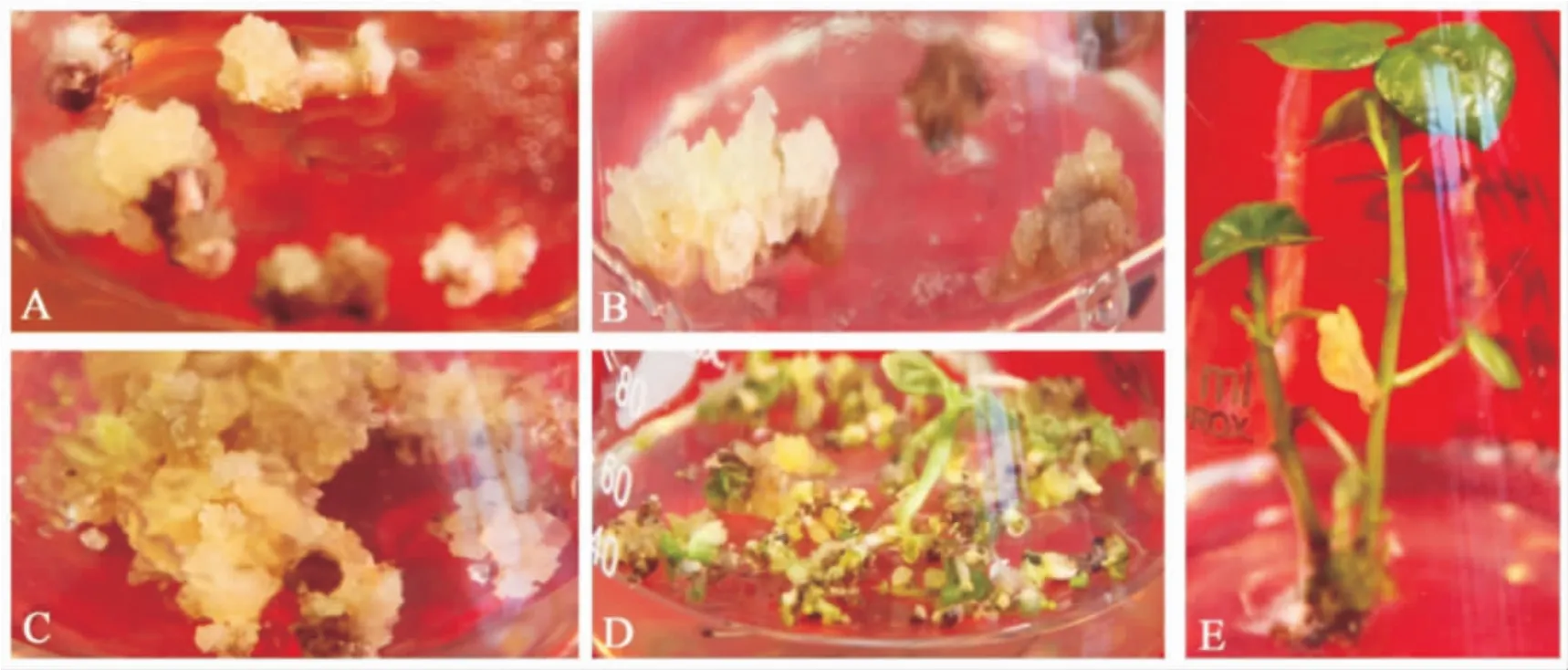
图3 转基因棉花的获得Fig.3 Development of transgenic cotton plants

图4 潮霉素涂抹叶片检测分析Fig.4 Phenotypes of leaves smeared with hygromycin

图5 部分转基因棉花株系的PCR检测结果Fig.5 The PCR detection of the transgenic plants
将这22株转基因材料的种子继续种植,获得T1转基因材料。在T1群体的苗期(3~4叶期)涂抹潮霉素进行标记基因检测和抗蚜鉴定,收获兼抗棉蚜和潮霉素的植株种子,结合分子检测连续二代筛选,最终选育出4个农艺性状较好的T3抗蚜株系 CZ1,CZ2,CZ3 和 CZ4。
2.2 转基因棉花植株Southern blot分析
对PCR阳性且能够正常开花授粉的22株转基因材料进行分子杂交检测,分析外源基因片段的插入情况。Southern blot分析结果显示有12株为单拷贝插入,6株为双拷贝,4株多拷贝。图6所示,CZ1、CZ3、CZ4 为单拷贝,CZ2 为 2 个拷贝。这些结果说明利用农杆菌介导转化棉花获得的转基因植株主要是单拷贝。
2.3 转基因棉花植株的转录表达分析
将收获种子的22个转基因植株种植在网室进行PCR检测筛选纯合转基因系,最后共获得农艺性状较好的纯合转基因系4个,分别是图6中的CZ1、CZ2、CZ3和 CZ4, 其中 CZ2为双拷贝插入。对这4个转基因株系进行转录表达分析,RT-PCR分析结果表明4个纯合系的外源双链RNA均被表达,能够扩增出目的条带,而对照没有扩增出相应的条带(图7)。 其中 CZ1、CZ2、CZ3中外源基因片段表达量相对较高,CZ4表达量相对较低。这些结果表明转基因棉花材料能够表达双链RNA。
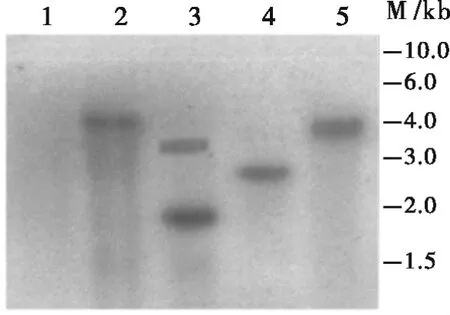
图6 部分转基因棉花株系的Southern杂交分析Fig.6 Southern blot analysis of the transgenic plants

图7 转基因棉花的AgApe基因片段表达分析Fig.7 The expression analysis of AgApe in transgenic cotton lines
2.4 转基因棉花的抗蚜性分析
大田的蚜虫抗性鉴定结果同网棚(表3)基本一致,4个转基因的株系的蚜害指数减退率极显著高于对照;图8显示在大田种植的这4个转基因材料和受体对照,对照叶片严重卷曲,而转基因植株受蚜虫危害情况明显低于对照植株。根据表2棉花蚜虫抗性鉴定分级标准,2015年的大田鉴定结果显示CZ1和CZ3为高抗棉蚜级别;且两年的蚜害指数减退率大田略高于网棚,可能是在大田自然状态下存在棉蚜天敌的原因。2016年网棚鉴定结果CZ4的蚜害指数减退率为42.5%,为中抗级别;2016年网棚和大田的的蚜害指数减退率均低于2015年,其中原因有待进一步的分析研究。
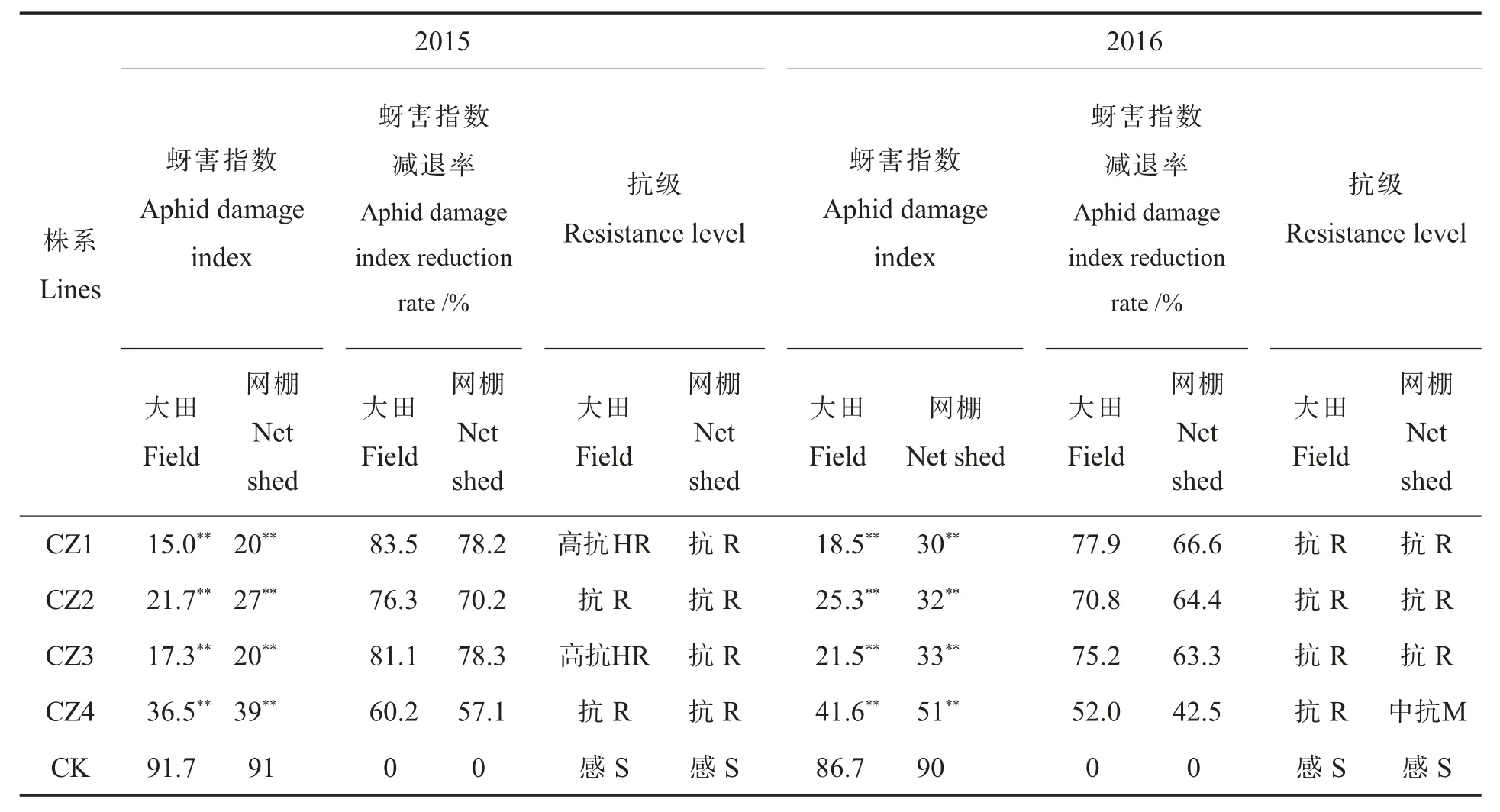
表3 转基因株系蚜虫抗性鉴定结果Table3 Results of resistance identification of transgenic plant aphids

图8 田间抗蚜虫表型Fig.8 Phenotypes of aphid resistance of transgenic lines and the control in field
3 讨论
近年来RNAi在植物虫害防治等方面研究报道日益增多。高朝宝等[22]构建了韧皮部特异性表达的RNAi干扰载体并将其转入到野生型拟南芥中,对转基因拟南芥接种棉蚜,抗蚜实验数据显示效果不明显,推测可能是因为实验过程中对蚜虫产生损害而导致实验数据误差以及实验过程不严谨造成的。熊叶辉[23]取棉铃虫蜕皮调节转录因子HaHR3为靶标基因,通过原核表达HaHR3的不同dsRNA片段筛选出目的靶标的干扰片段。将此片段构建转基因植物表达干扰载体,通过转基因植物介导的RNAi沉默棉铃虫HaHR3基因表达,棉铃虫取食含有dsRNAs的饲料和转基因植物后,HaHR3基因的转录水平及蛋白表达水平均被有效抑制,导致了棉铃虫幼虫的死亡及生长发育畸形。耿文超[24]在棉叶螨TtAK与TtCHI双价RNAi载体的构建及抗虫性初步研究中,对转基因烟草与野生型烟草上进行接虫实验,结果表明:抑制TtAK和TtCHI双基因的转基因烟草显著降低了叶螨的存活率,产卵量减少约23%~30%,叶螨生育周期缩短约4 d,但对虫卵的孵化率并没有影响。目前RNAi在棉花抗蚜性研究上还尚未见报道。
棉蚜一直是我国各棉区的主要害虫之一,也是棉花害虫防治的重点。随着转Bt基因抗棉铃虫棉花的成功研制和广泛种植,棉铃虫危害得到了有效的控制,而棉蚜已经上升为重点防治对象。我们利用棉蚜的腺苷三磷酸酶E亚基基因构建RNAi载体,并转化中棉所35,转基因棉花对棉蚜有较高的抗性,为棉花抗蚜育种提供了新思路和新方法。
4个转基因纯合系材料表达分析结果显示,CZ1、CZ2、CZ3中转基因片段的表达量相对较高,CZ4表达量相对较低,CZ4植株顶部由于蚜虫危害叶片有卷曲,而其他3个转基因材料CZ1、CZ2、CZ3植株顶部叶片平展(图8),不同株系间抗性差异除了与基因的插入事件相关外,也可能与植物双链RNA容易被加工成siRNA有关[25]。根据棉花蚜虫抗性鉴定分级标准,4个转基因株系的蚜害指数减退率极显著高于对照,网棚蚜虫抗性鉴定结果和大田蚜虫抗性鉴定结果基本一致。2016年的蚜害指数减退率低于2015年,其中原因有待进一步的分析研究。
RNAi技术是棉花抗蚜基因育种的1个新途径,由于棉蚜mRNA具有特异性,相应的dsRNA或者siRNA也具有特异性,只作用于棉蚜而不伤害棉田其他生物,不作用于非靶标生物或天敌昆虫,没有生态风险。外源基因片段转入棉花植株后,转基因棉株能够正常的开花结籽,通过连续二代筛选证明,外源基因片段可以稳定地遗传给后代,因此利用RNAi技术防治棉蚜的本项研究结果为培育抗蚜棉花新品种看到了希望。
4 结论
本研究利用蚜虫腺苷三磷酸酶E亚基基因,构建其RNAi载体,利用农杆菌介导法转化棉花。通过PCR检测获得22株阳性转基因株;分子杂交鉴定发现了12个转基因株为单拷贝插入。结合田间农艺性状和dsRNA表达量,我们筛选出4个转基因株系:CZ1、CZ2、CZ3、和 CZ4。通过 2 年的网棚和大田的蚜虫抗性鉴定,结果表明转基因棉花CZ1、CZ2和CZ3具有较高的抗蚜性,两年的蚜害减退率均在60%以上。利用RNAi技术获得了高抗棉蚜的棉花材料,为棉花抗蚜育种提供了新种质。
猜你喜欢
当代水产(2022年7期)2022-09-20
河北科技师范学院学报(2022年2期)2022-08-26
小天使·一年级语数英综合(2022年5期)2022-05-25
中国饲料(2021年17期)2021-11-02
发明与创新·小学生(2021年5期)2021-08-23
空间科学学报(2021年1期)2021-05-22
发明与创新(2021年19期)2021-05-07
环境昆虫学报(2021年1期)2021-03-30
农民致富之友(2020年33期)2020-12-03
新疆农业科学(2020年11期)2020-10-27
