
MPTP模型小鼠中Th17及其炎症因子的作用研究
2018-10-29 09:23:38方杰吕哲刘嘉音王琛洪甘济郑良城马琪林
中国现代药物应用 2018年19期
方杰 吕哲 刘嘉音 王琛 洪甘济 郑良城 马琪林
帕金森病(Parkinson disease, PD)是仅次于阿尔兹海默病的世界第二大退行性疾病[1]。其主要病理特点为黑质纹状体多巴胺(dopamine, DA)能神经元选择性、渐进性的变性缺失, 胶质细胞增生和细胞胞内路易氏小体(Lewy body)形成[2]。病因机制仍不完全清楚, 其中免疫炎症机制在其发病过程中起着重要的作用[3]。CD4++Th淋巴细胞亚群Th17细胞参与了许多中枢神经系统疾病的发生和发展[4], 在PD模型中,N-α-synuclein可刺激Th17相关基因的高表达, 诱导Th17分化, 促进炎症级联反应, 从而介导多巴胺能神经元的损伤[5]。本研究应用MPTP小鼠模型, 研究Th17特异性转录因子RORγt和特异性炎症因子IL-17对PD的作用, 进一步明确Th17在PD中的作用机制。现报告如下。
1 材料与方法
1.1 实验动物 选择14~15周龄、雄性、体重(27±2)g的C57BL/6小鼠, 常规饲养于厦门大学实验动物中心。
1.2 实验方法
1.2.1 PD模型构建 将48只C57BL/6小鼠随机分为PD模型组和对照组, 每组24只。PD模型组:造模第1天按照MPTP 20 mg/(kg·次), 每次间隔2 h, 腹腔注射4次。对照组:同样的方法按照0.05 ml/次注射生理盐水。两组小鼠均在造模第2天开始, 连续腹腔注射生理盐水7 d, 0.2 ml/(次·d)。
1.2.2 行为学检测 小鼠行为学检测转棒仪速度设置为5 min内从4 r/min逐渐增加到40 r/min, 每次检测间隔5 min,连续测3次取平均值。
1.2.3 中脑黑质TH免疫化学染色 实验小鼠进行行为学检测后, 每组各取小鼠6只进行取材。取出中脑常规处理后以12 μm冠状位连续冰冻切片, 室温封闭30 min, 1∶200比例稀释的抗TH兔多克隆抗体覆盖, 孵育48 h。再次洗涤后将1∶200比例稀释的FITC标记的山羊抗兔免疫球蛋白G(IgG)抗体滴入切片上, 孵育6 h。洗涤后封片, 后将片子荧光正置显微镜下观察, 计数每只小鼠10张切片的TH阳性神经元数目。
1.2.4 酶联免疫吸附实验(ELISA) 法检测外周血IL-17的表达 行为学实验后的小鼠每组取6只常规处理后进行眼球取血制备血清样本。复孔加入100 μl梯度稀释的标准品, 样本孔加入20 μl的样本和80 μl的1×检测缓冲液, 并在每孔中加入50 μl稀释后的检测抗体, 室温下孵育1.5 h。洗涤后每个微孔中加入100 μl稀释后的辣根过氧化物酶标记的链霉亲和素, 孵育30 min。再次洗涤后每孔加入100 μl的底色显物TMB, 室温下避光反应20 min, 测定样本光密度(OD)值, 并根据标准曲线计算样品中检测因子的含量。
1.2.5 Real-time聚合酶链式反应(PCR)法检测中脑黑质中RORγt和IL-17的mRNA表达水平 行为学实验后每组取6只小鼠常规处理取中脑黑质组织, Trizol提取总RNA, 逆转录合成cDNA, Primer 5设计引物, PCR反应条件设置为:95℃预变性30 s, 95℃变性5 s, 60℃退火30 s, 72℃延伸20 s,40个循环。记录内参基因与目的基因的Ct值, 运用2-△△Ct法进行数据分析。
1.2.6 Western blot法检测中脑黑质中RORγt和IL-17的蛋白表达水平 行为学实验后的小鼠每组取6只常规处理取中脑黑质组织制备样本, 聚丙烯酰胺凝胶电泳分离、电转。电转过后的聚偏二氟乙烯(PVDF)膜用TBS/T洗膜液清洗,封闭2 h后加入一抗溶液, 4℃包被过夜。洗涤后加入对应的辣根过氧化物酶标记的二抗溶液, 室温孵育1 h。再次洗涤后的PVDF膜用增强型化学发光剂溶液浸润, 于全自动化学发光仪曝光。
1.3 统计学方法 采用Graphpad Prism 5软件对研究数据进行统计分析。计量资料以均数±标准差()表示, 采用独立样本t检验和单因素方差检验。P<0.05表示差异具有统计学意义。
2 结果
2.1 行为学检测及免疫荧光染色 PD模型表现出肢体震颤、尾部僵直、运动减少等异常行为学反应, PD模型组转棒时间为(17.6±2.3)s显著短于对照组的(112.1±8.6)s, 差异具有统计学意义(P<0.05)。见图1。PD模型组中脑TH阳性反应的神经元数目较对照组明显减少。见图2。
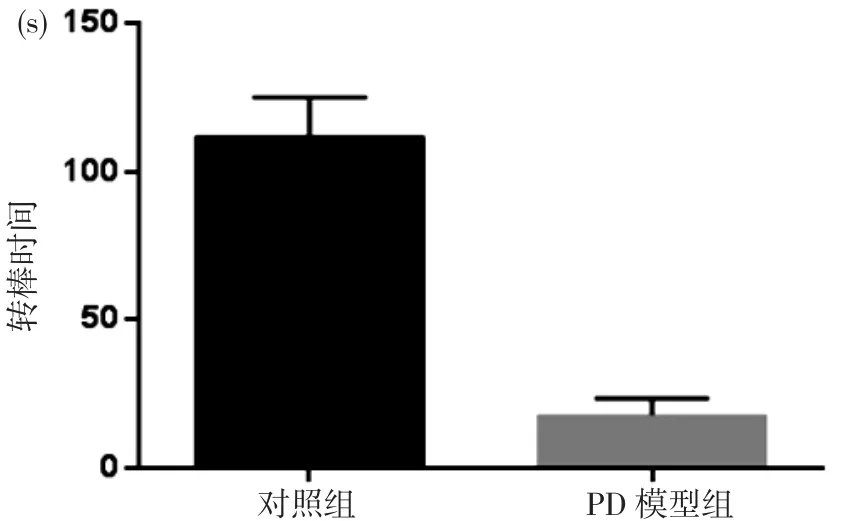
图1 转棒行为学实验两组小鼠转棒时间

图2 两组小鼠中脑TH阳性反应神经元免疫荧光染色图
2.2 IL-17和RORγt的表达水平 PD模型组外周血IL-17表达为(32.58±6.21)高于对照组的(7.46±2.10), 差异有统计学意义(P<0.05)。见图3。PD模型组中脑黑质中RORγt和IL-17的mRNA表达水平分别为(8.87±1.12)、(5.66±0.85)高于对照组的(2.64±0.52)、(2.08±0.41), 差异有统计学意义(P<0.05)。见图 4, 图 5。

图3 两组小鼠外周血IL-17表达水平

图4 两组小鼠中脑黑质中IL-17和RORγt的mRNA表达水平

图5 两组小鼠中脑黑质中IL-17和RORγt的蛋白表达水平的免疫印迹图
3 讨论
免疫炎症是PD的重要病理机制[3], 抗炎药物降低罹患PD的风险提示了进一步研究炎症机制及寻找可能的治疗靶点具有重要意义[6]。本实验中观察到PD模型出现躯体僵直、震颤、竖毛、运动减少等行为异常表现。转棒仪检测显示PD模型组转棒时间显著短于对照组, 差异具有统计学意义(P<0.05)。中脑组织TH特异性染色显示PD模型组中脑DA能神经元显著少于对照组。这些表明MPTP可诱导PD样病理损害[7]。
淋巴细胞可穿过血脑屏障进入中枢神经系统[8], 在PD病程中, CD4++T细胞在神经毒性损伤机制中发挥着重要作用[9]。Th17细胞是新近发现的CD4++T细胞, 有研究表明Th17是介导MPTP模型小鼠DA能神经元损伤的主要效应细胞[5]。本研究显示PD模型外周血中IL-17显著高表达, 证明系统性免疫炎症反应可能参与其致病机制。PD模型的中脑黑质中RORγt和IL-17在基因和蛋白水平一致性表达上调, 表明Th17细胞及其炎症因子可能参与了PD的中枢神经系统病理损伤过程。结合既往研究结果, 推测Th17引起的系统性免疫炎症反应和中枢神经系统免疫炎症反应可能协同促进了PD的病理损害。
综上所述, Th17细胞及其炎症因子可能通过诱发外周和中枢神经系统免疫炎症反应参与PD的发生及发展, 进一步深入研究其具体作用和效应机制, 为探索PD免疫治疗新方法奠定基础。
猜你喜欢
浙江医学(2020年9期)2020-07-01 10:17:42
中成药(2017年9期)2017-12-19 13:34:27
CHINESE JOURNAL OF AERONAUTICS(2017年1期)2017-11-21 12:54:14
中成药(2017年5期)2017-06-13 13:01:12
卒中与神经疾病(2017年1期)2017-01-11 10:12:50
中国医疗器械杂志(2015年5期)2015-12-31 06:15:20
磁共振成像(2015年7期)2015-12-23 08:53:04
医学研究杂志(2015年6期)2015-07-01 17:40:53
环球中医药(2015年4期)2015-02-27 15:01:51
西南军医(2015年5期)2015-01-23 01:25:00
