
海州露天矿排土场不同复垦模式下土壤酶与土壤肥力典型相关分析
2018-10-22 03:00:26刘宝勇宋子岭李岩岩
中国水土保持科学 2018年4期
刘宝勇,赵 凯,宋子岭,李岩岩
(辽宁工程技术大学,123000,辽宁阜新)
土壤酶对生态系统的物质转化、能量流动以及土壤肥力的形成起着至关重要的作用,土壤中几乎所有的生物和化学物质都依赖于酶的催化[1-2],且与土壤理化性质和环境状况关系密切,是土壤生物指标研究中优先考虑的指标之一[34],由于土壤酶的敏感性,已经成为了解土壤状况的指示性指标之一[56]。周礼恺[7]和方瑛等[8]就不同样地土壤酶活性与土壤肥力的关系进行了研究,并认为土壤酶可以作为评价土壤肥力的指标。
国外的复垦工作研究比国内早,并且主要的研究内容集中在土壤重构和基本特性方面[910]。而我国初步建立了矿区塌陷区、露天矿排土场等废弃地复垦土壤的原理和方法,并对水土流失做了研究[1114]。土壤酶活性与土壤肥力因子的关系研究多以粮油作物田地土壤为研究对象,以排土场复垦土壤为研究对象较少,而且多应用相关分析和通径分析研究土壤酶活性与土壤性质的关系[1516]。对露天矿排土场复垦土壤的研究主要集中在复垦模式对土壤酶活性[17]、土壤理化性质、微生物群落及土壤养分评价体系的建立[1819]等方面。上述研究对解释海州露天矿排土场土壤生物活性及养分状况提供了依据,鲜有对排土场复垦土壤酶与土壤肥力相关性方面进行研究,闫晗等[17]研究了海州露天矿排土场6种土地利用方式对土壤养分及土壤酶活性的影响,发现人工修复地和天然草地可以显著提高土壤养分和土壤酶活性。之前研究大都讨论单因子间的相关性,本研究以典型相关分析为主,研究2组变量之间相关性,揭示2组变量之间的线性相关关系[2022],典型相关分析比一般相关分析更加全面地反映变量之间的内在联系[23]。
为了进一步改善排土场的水土流失状况,笔者选择排土场中的刺槐(Robinia pseudoacacia L.)林地、白榆(Ulmus pumila L.)林地以及草地3种复垦模式为研究对象,以未复垦的荒裸地作为对照,探究复垦后的土壤酶活性(碱性磷酸酶、蔗糖酶、过氧化氢酶、蛋白酶、脱氢酶以及脲酶)以及土壤肥力因子(有机质、全氮、全磷、速效钾)和pH值等土壤理化性质的相关性,对排土场的生态恢复及重建具有指导意义。
1 研究区概况
研究区为辽宁省阜新市海州露天矿排土场,该地区年平均降水量539 mm,降水集中于7—9月,年平均蒸发量达1 800 mm,年平均气温7.3℃,昼夜温差大,相对湿度50% ~60%,是典型的北温带半干旱大陆性季风气候区。试验样地为复垦年限20年的西排土场,其土质是由露天矿排出的页岩、砂岩构成[18]。
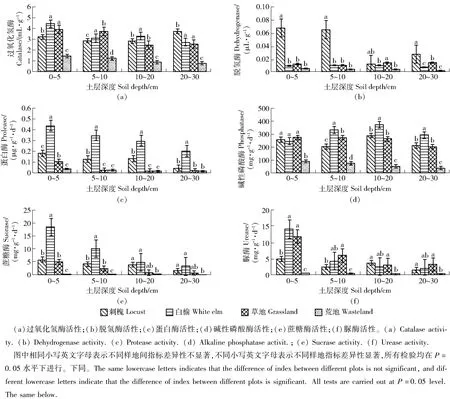
图1 不同复垦模式土壤酶活性Fig.1 Soil enzyme activities in different reclamation modes
2 研究方法
按照混合法采集0~30 cm深的土壤,分0~5、5~10、10~20和20~30 cm 4层。每1层任意取4个样点,同层混合。其中在刺槐林地和白榆林地划出10 m×10 m的标准地,在标准地中通过测量胸径选择标准木,在距离标准木根部0.75 m的圆周上平均分布4点进行采样,同层均匀混合。而荒草地和荒裸地(对照)则划出一块长势较为平均的10 m×10 m的样方,在样方内采用“S”法进行采样。
过氧化氢酶活性采用容量法测定[24],脲酶活性采用次氯酸钠比色法测定[24],脱氢酶活性采用TTC比色法测定[24],蛋白酶活性采用茚三酮比色法测定[24],蔗糖酶活性采用二硝基水杨酸比色法测定[24],碱性磷酸酶活性采用磷酸苯二钠比色法测定[24],全氮采用全自动凯氏定氮法测定,全磷采用NaOH 熔融 钼锑抗比色法,速效钾采用火焰分光光度计法,有机质采用重铬酸钾氧化 外加热法测定,土壤pH值采用pH计测定[25](水土比2.5:1),具体操作方法参照《土壤农业化学分析方法》[25]。
用Excel 2007进行做图分析以及SPSS16.0软件对所得数据进行单因素方差分析(One-way ANOVA)及典型相关分析[26]。
3 结果分析
3.1 不同复垦模式下土壤酶活性及土壤肥力分析
3.1.1 土壤酶活性分析 由图1可见,不同样地土壤酶活性表现出显著的差异性(P<0.05),刺槐林地、白榆林地和草地土壤酶的活性均显著高于荒裸地。从图1(a)可知:过氧化氢酶促进过氧化氢的分解有利于防止其对生物体的毒害作用,过氧化氢酶活性与土壤有机质含量有关[27],在垂直方向上,刺槐林地过氧化氢酶活性在20~30 cm土层最高,是表层土壤的1.16倍,这与该层土壤有机质含量高有关;白榆林地与草地均为表层过氧化氢酶活性最高,活性脱氢酶是土壤生物细胞的一部分[28],因此测量脱氢酶活性实际上是代表土壤微生物的瞬时代谢活动[29]。从图1(b)可知:0~5和5~10 cm土层脱氢酶变化规律几乎一致,均为刺槐林地酶活性最高,其他样地无显著性差异;草地在总体上土壤酶活性高于白榆林地,尤其在20~30 cm土层,草地脱氢酶活性是白榆林地的2倍。从图1(c)可知:蛋白酶参与土壤中氨基酸、蛋白质以及其他含蛋白质氮的有机化合物的转化,它们的水解产物是高等植物的氮源之一,蛋白酶活性均呈现出随着剖面深度而减弱的趋势,白榆林地蛋白酶活性最高,因为白榆为固氮树种;除了0~5 cm土层蛋白酶活性草地>荒裸地,其他土层差异性不显著,可能因为表层土壤草地根系分布密集,受到动物或微生物活动的影响。从图1(d)可知:碱性磷酸酶活性在5~30 cm土层均为白榆林地活性最高,0~5 cm草地酶活性最高,刺槐、白榆、草地酶活性相差无几,荒裸地表层土壤酶活性最高,并随着土层加深,酶活性递减。从图1(e)可知:蔗糖酶的变化规律与蛋白酶十分类似,这是因为土壤剖面中蔗糖酶的分布规律与蛋白酶类似。各个样地蔗糖酶活性均为表层酶活性最高,随着土层深度的加深,酶活性递减,其中白榆林地酶活性最高。从图1(f)可知:尿素水解后的某些酶统称为脲酶,其酶促反应的主要产物是氨气,因此脲酶与土壤中的氮关系密切,在0~5 cm土层,除了荒裸地外的各个样地脲酶活性显著高于其他土层,可能因为土壤表层覆盖枯枝落叶,微生物活性较高,酶促底物较荒裸地多;5~10 cm草地土壤受到根系影响,所以脲酶含量最高。
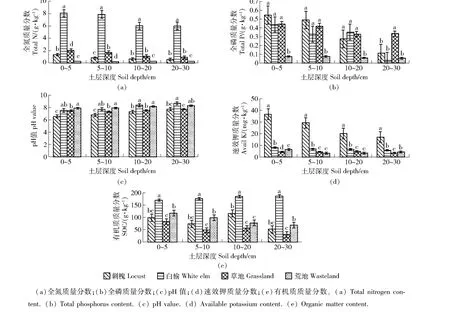
图2 不同复垦模式土壤肥力Fig.2 Soil fertility of different reclamation modes
3.1.2 土壤肥力分析 从图2看出:本研究所选取的4种样地土壤pH值均>7,偏碱性,全氮、全磷、速效钾、有机质质量分数以及pH值之间具有显著性差异。如图2(a)所示:氮素是影响土壤肥力的重要指标,不同复垦模式土壤氮素含量不同,白榆林地全氮质量分数显著高于其他样地,随着土层的加深,土壤全氮质量分数呈现出不同程度的递减趋势。如图2(b)所示:刺槐林地、草地及荒裸地全磷质量分数随着土层深度增加,质量分数呈递减趋势;而白榆林地则出现先减后增再减的情况;除了20~30 cm土层,刺槐、白榆和草地的全磷质量分数均无显著性差异。如图2(c)所示:土壤pH值是土壤的重要化学性质,复垦地的pH值较未复垦地有所降低,使pH值大致为中性,并且随着土层加深,pH值增加,但变化不显著。如图2(d)所示:在各个土层中,刺槐林地速效钾质量分数显著高于其他样地;刺槐林地较白榆林地、草地和荒裸地速效钾质量分数分别提高了283.9%、428.8%和503.1%。如图2(e)所示:白榆林地和荒裸地的有机质质量分数较高,因为蔗糖酶活性与碱性磷酸酶活性有助于提高有机质质量分数[12];并且随着土层深度的增加,土壤有机质质量分数呈递增的趋势,但变化不显著。荒裸地随着土层深度的加深,土壤有机质质量分数呈逐渐递减。土壤有机质质量分数依次为白榆林地>荒裸地>刺槐林地>草地。
3.2 土壤酶活性与土壤肥力的典型相关分析
按照典型相关分析原理,土壤养分因子数据是X(5×16),土壤酶活性因子数据应为Y(6×16)。在分析过程中,层面缩减度分析1TO5、2TO5和3TO5的典型相关系数<0.05,达到显著水平,故选取3组变量(U为土壤肥力因子,V为土壤酶活性)来表示X组和Y组之间的相关关系。由表1可见:典型向量1、2、3的典型相关系数具有统计学意义,说明土壤酶活性及土壤肥力可以由前3对典型变量进行描述。并且本实验中各个变量的单位不统一,需要对数据进行标准化处理,所以选用因变量的标准化典型加权系数。
第1组典型变量为:


式中:X1为全氮质量分数,g/kg;X2为全磷质量分数。g/kg;X3为速效钾质量分数,mg/kg;X4为有机质质量分数,g/kg;X5为pH值。Y1为过氧化氢酶,mL/g;Y2脱氢酶,μL/g;Y3碱性磷酸酶,mg/(g·d);Y4为蔗糖酶,mg/(g·d);Y5为蛋白酶,μg/(g·d);Y6为脲酶,mg/(g·d)。
由表2可见,X的变异被U1(自身典型变量)解释的比例是35.29%,Y的变异被V1(自身典型变量)解释的比例是49.25%,说明X、Y的信息主要集中在第1对典型变量上。酶活性指标Y的变异被U1解释的比例是49.57%,X的变异被V1解释的比例是35.06%。X的变异被 U2解释的比例是40.77%,Y的变异被V2解释的比例是10.93%;X的变异被V2解释的比例是39.61%,Y的变异被U2解释的比例是11.25%,说明第2对典型变量受土壤肥力指标X的影响较大。综合第3对典型变量,土壤酶活性指标影响大于土壤肥力指标。
经分析得出,第1组典型变量的相关系数为0.997(表2),经检验相关变量呈极显著相关关系。综合土壤肥力因子系数较高载荷较大的是全氮(X1)和速效钾(X3),其次是全磷(X2);综合土壤酶活性因子载荷大的是蛋白酶(Y5)。土壤蛋白酶活性与肥力指标全氮、全磷和速效钾质量分数关系密切。第2组典型变量相关系数为0.986(表2),经检验相关变量呈显著相关关系。综合土壤肥力因子载荷最大的是有机质(X4),其次是全氮(X1)、全磷(X2)、和pH值(X5);综合土壤酶活性因子载荷较大的是蔗糖酶(Y4)和蛋白酶(Y5)。蔗糖酶及蛋白酶活性与全氮、全磷、有机质和pH值关系密切。第3组典型变量相关系数为0.871(表2),经检验典型变量呈相关关系,综合土壤肥力因子载荷最大的是土壤pH值;综合土壤酶活性因子载荷最大的是蔗糖酶(Y4),其次是蛋白酶(Y5)和脲酶(Y6)。蔗糖酶活性、蛋白酶活性、脲酶活性与pH值的关系密切。
4 结论与讨论
4.1 不同复垦模式比较
不同复垦模式下土壤酶活性以及土壤肥力均存在一定差异性。结果表明,白榆林地除了速效钾与脱氢酶,土壤酶活性与土壤肥力的各项指标均较高,且较其他复垦模式差异性显著。在各个土层中,白榆林地蔗糖酶活性与蛋白酶活性显著高于其他样地(P<0.05)。在0~5 cm土层中,草地土壤过氧化氢酶活性、碱性磷酸酶活性和脲酶活性均优于刺槐,这是因为表层土壤各种草根系密集,受根系分泌物影响,各种酶活性指标良好。在各层土壤中,刺槐和白榆林地蔗糖酶活性提升最为显著,分别较荒裸地提升了24.8倍和77.8倍。而在20~30 cm土层中,刺槐林地土壤酶活性则优于草地,这是因为草根系分布锐减,而刺槐毛根根系的分布则会增加。草地则对脲酶活性提升较为显著,提升了28.1倍,这与闫晗等[17]的研究结果相一致,这是因为白榆为固氮树种,根系有较强的固氮作用,白榆林地土壤中全氮和有机质质量分数都相对较高。有所差异的是人工种植的白榆在提高土壤蛋白酶的能力要高于刺槐,土壤蛋白酶活性、蔗糖酶活性和脱氢酶活性随着土层的加深,酶活性有递减的趋势,表层土壤的酶活性最高,这与枯落物有关[30],土壤酶可以促进枯落物分解,促进地表微生物的活性,提高了土壤酶活性。土壤过氧化氢酶活性、碱性磷酸酶活性和脲酶活性未表现出明显的规律性,甚至在20~30 cm土层部分土壤酶的活性会出现跃升,而李智兰[31]的研究却发现随着土层加深,土壤脱氢酶活性、碱性磷酸酶活性以及脲酶活性总体呈递减的趋势,造成这种差异可能是由于排土场深层土壤粉砂岩、砾岩以及煤质岩含量的陡然增加,从而间接地影响了土壤酶活性。刺槐林地0~5 cm土层脱氢酶活性显著高于其他林地,可能是由于受到外界因素的干扰,如放牧或矿质残渣等原因[32],长期使用粪肥和矿肥能够显著提高土壤酶活性[33]。各个土层白榆林地土壤全氮和有机质显著高于其他样地(P<0.05)。全氮能够促进植物根际微生物生长,有利于植物养分吸收以及土壤酶合成[34]。除了pH值外,土壤肥力因子随着土层深度的增加,土壤肥力有所减少,表现出了“表聚效应”,这与张丽娟等[11]研究成果一致,这与表层土壤的枯落物和植物根系分布有相关性。复垦地土壤pH值低于未复垦地,复垦地土壤pH更接近于中性,并且随着土层深度的增加pH值有所升高,但变化不显著,并且刺槐稳定土壤pH效果更好,与王翔等[27]的研究结果一致。pH值不仅影响有机大分子分解与矿化、微生物种类及活性等,而且影响土壤酶酶促反应速率。综上所述,白榆林地的复垦效果好于其他林地。
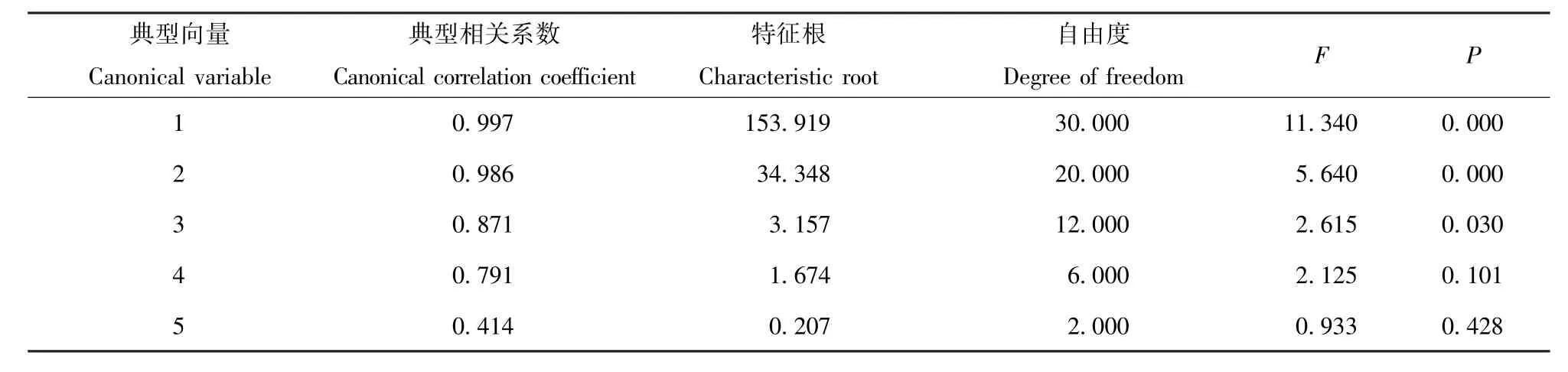
表1 典型变量的显著性检验Tab.1 Significance test of canonical variables(n=16)

表2 变量的变异能由典型变量解释的比例Tab.2 Proportion of variation in variables that can be explained by canonical variables(n=16)
4.2 蔗糖酶、蛋白酶和脲酶与土壤肥力相关性显著
通过典型相关分析研究发现:综合土壤酶活性因子中其主要作用的是蛋白酶、蔗糖酶和脲酶;综合土壤肥力因子中所有因子都参与其中,说明蛋白酶、蔗糖酶和脲酶参与了土壤C、N、P的转化以及对土壤pH值的调控也起到一定的作用,与安韶山等[26]研究丘陵区土壤发现蔗糖酶活性与土壤肥力因子有显著的相关性的结果相一致。蔗糖酶是评价土壤恢复过程中的重要指标,并且有机质的分解深刻地影响着土壤酶的活性[35]。并且K.Ciarkowska等[36]通过研究铅锌矿土壤修复过程中酶活性发现土壤样品中的有机质和氮的高可变性反映在脲酶和蔗糖酶的变化上。K.Ciarkowska等[36]还发现土壤有机质和蔗糖酶活性可以作为评价复垦工程的间接指标。过氧化氢酶、脱氢酶和碱性磷酸酶与土壤肥力的关系均不显著,这与李跃林等[20]在桉树人工林地中研究发现土壤过氧化氢酶活性、脲酶活性和蛋白酶活性与土壤肥力因子有显著相关性的结果有所差异。本研究中未见过氧化氢酶与土壤肥力有显著的相关性,这是因为试验地常年受到放牧等人为因素的干扰[37],土壤肥力因子影响分泌土壤酶的土壤微生物和植物根系的活动[38]。
以上分析结果可以发现露天矿排土场由于其特殊的立地条件,与其他研究区域不同,其土壤酶与土壤肥力因子有独特的相关性。因为蔗糖酶、蛋白酶和脲酶与土壤肥力因子有显著的相关性,所以在实际工作中,可以通过测定蔗糖酶和蛋白酶的活性快速评价露天矿排土场土壤肥力状况。
猜你喜欢
采矿技术(2021年5期)2021-10-12 08:24:22
现代园艺(2020年2期)2020-12-22 05:57:04
生物技术通报(2020年12期)2020-12-21 09:20:00
世界有色金属(2020年21期)2020-12-08 12:41:33
腐植酸(2020年1期)2020-11-29 00:15:44
中国设备工程(2019年17期)2019-09-24 09:25:14
小溪流(故事作文)(2018年10期)2018-12-28 10:45:08
浙江理工大学学报(自然科学版)(2018年6期)2018-11-07 11:14:32
现代园艺(2018年2期)2018-03-15 08:00:12
山东林业科技(2016年5期)2016-07-05 00:43:00
