
基于RIL群体的烤烟烘烤特性遗传分析
2018-10-19 10:04张玉琴王传义李青山刘要旭徐秀红
西南农业学报 2018年9期
张玉琴,孙 阳,王传义,李青山,刘要旭,徐秀红*
(1. 中国农业科学院烟草研究所,农业部烟草生物学与加工重点实验室,山东 青岛 266101;2. 中国农业科学院研究生院,北京 100081)
【研究意义】烘烤特性是指烟叶在烘烤过程中表现出的变黄和脱水的速度及同步程度,变黄后是否容易定色等特性,包括易烤性和耐烤性[1]。容易变黄,变黄和脱水协调的烟叶易烤性好,反之易烤性差。叶片变黄快且维持不变褐的时间越长,其耐烤性越好,烘烤特性越好[2]。研究烤烟烘烤特性的遗传基础可以为选育烘烤特性好的品种和提高烤烟烘烤品质提供理论基础,对烟草品质育种具有一定的指导意义。【前人研究进展】烟叶烘烤从外观看是叶绿素等色素降解的反映,藤田茂隆等[3]以单倍体加倍系为研究材料,提出用变黄指数来指示烟叶的易变黄性,揭示了烤烟易烤性很可能受一个主效基因控制,并发现易烤性与生物碱含量密切相关。另有研究[4-5]表明烤烟耐烤性与多酚氧化酶活性密切相关,多酚氧化酶的差异特性主要取决于品种和栽培条件。性状间的连锁证明烟叶烘烤特性受遗传影响[3,6-7]。近几年国外对烤烟烘烤特性的研究报道较少,国内对烘烤特性的遗传研究有所加强。中国农业科学院烟草研究所利用烤烟组合的四世代(P1、P2、F1、F2)及六世代群体(P1、P2、F1、F2、B1、B2),初步揭示了烤烟烘烤特性的遗传规律,用SSR分子标记构建遗传图谱,初步定位了与烘烤特性相关的QTL位点[8-10]。D.Gong等[11]结合SLAF-Seq开发的SNP标记与公布的烟草基因组SSR分子标记[12-13]构建烤烟分子遗传连锁图谱,发现了2个易烤性主效QTL并将其定位到了烟草基因组的第6个连锁群上。
植物数量性状的主基因+多基因混合遗传模型广泛应用于水稻[14]、小麦[15]、玉 米[16]、黄瓜[17]、棉花[18]、烟草[19-21]等粮食、蔬菜和经济作物。利用重组自交系对烤烟烘烤特性进行遗传分析的研究尚未有报道。【本研究切入点】本研究以烤烟品种云烟85和大白筋599构建的重组自交系群体为试验材料,利用“主基因+多基因”的植物数量性状遗传分析方法,对烤烟易烤性、耐烤性的遗传规律进行研究。【拟解决的关键问题】为烤烟烘烤特性QTL精细定位及应用提供参考。
1 材料与方法
1.1 试验材料
试验采用烘烤特性好的烤烟品种云烟85作为母本(P1),烘烤特性差的大白筋599作为父本(P2),从F1单株上收取自交种子,单粒传法获得F2∶7代重组自交系。
1.2 试验方法
1.2.1 试验设计 2016年在中国农业科学院烟草研究所青岛试验基地种植168个F2∶7代重组自交系。试验地土壤类型为棕壤,偏砂质土。年平均降雨量750 mm,年平均气温12 ℃。试验地土壤肥力状况:碱解氮59.45 mg/kg,有效磷23.60 mg/kg,速效钾106.33 mg/kg,有机质含量0.97 %,pH5.29,氯离子6.80 mg/kg。烟田行距1.2 m,株距0.5 m,留叶数20片,按照优质烟生产要求进行田间管理。试验按照完全随机区组设计,设3个重复。RIL家系每小区种植10株,共30株,亲本每小区种植20株,共60株。待到中部烟叶(10~11叶位)成熟后,RIL家系的每个重复取5片调查烘烤特性,亲本的每个重复取10片调查烘烤特性。
1.2.2 烘烤特性调查方法 采用烟叶成熟期暗箱试验[1]作为烘烤特性的测定指标。待烟叶成熟后,将其挂在保持室温的暗箱中,每24 h记录1次变黄比例和变褐比例,累计测9次。取24、48、72、96 h的变黄面积计算变黄指数(YI),YI=ΣY/n,Y表示每次的黄色面积比例,n表示测量次数。变黄指数越大,表示易烤性越好。从开始变褐进行变褐面积统计,变褐指数BI=ΣB/n,B表示变褐指数,n表示测量次数。变褐指数越小,表示耐烤性越好。
1.3 数据分析
方差分析采用SPSS 23.0,频率分布和曲线拟合采用Origin软件。遗传数据采用主基因+多基因的植物数量性状混合遗传模型分析。
2 结果与分析
2.1 亲本及RIL群体烘烤特性的表型结果
2.1.1 烘烤特性指标的方差分析 从表1可知,变黄指数在亲本间存在高度统计学意义,在亲本内重复间无统计学意义(P1=0.9065,P2=0.9174)。变褐指数在亲本间有高度统计学意义,在亲本内重复间无统计学意义(P1=0.9922,P2=0.8696),说明亲本在易烤性、耐烤性性状上存在真实有效的遗传差异。偏度和峰度的绝对值均少于1,说明变黄指数和变褐指数在亲本内的分布均服从正态分布,符合数量性状的分布要求[22]。RIL家系间变黄和变褐指数均具有高度统计学意义(P=0.0001),说明烤烟烘烤特性在家系间存在稳定的遗传差异,可以进行数量性状的遗传分析。

表1 父母本及重组自交系烘烤特性指标分析
注:同行标有A、B者表示亲本间差异有高度统计学意义(P<0.01)。
Note:The same lines label A and B indicate a statistically significant difference between the parents (P<0.01).
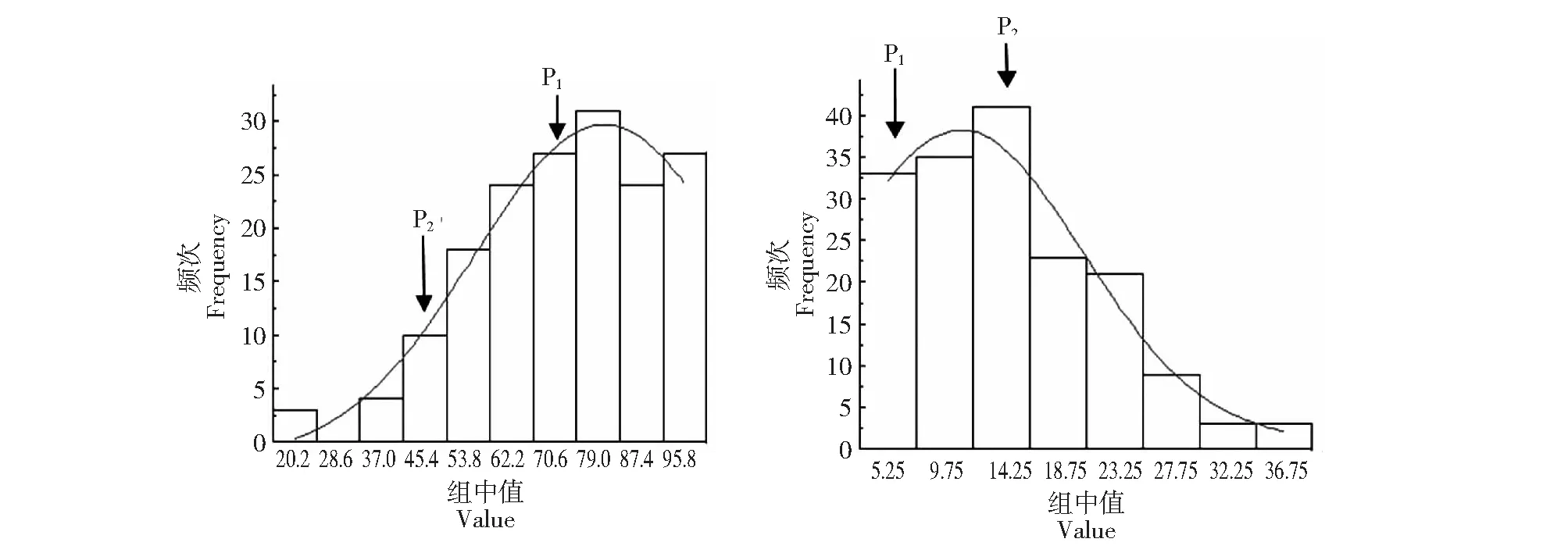
Y:变黄指数;B:变褐指数图1 变黄指数、变褐指数在重组自交系群体中的次数分布Fig.1 Frequency distribution of yellow index and browning index in RILs population
2.1.2 RIL群体烘烤特性指标的频率分布 由图1可以看出变黄指数和变褐指数在RIL群体中呈现连续分离,且均出现超亲现象,说明烘烤特性受多基因遗传或基因互作的影响。拟合曲线呈现单峰明显的偏态分布,说明易烤性、耐烤性的遗传可能受主基因控制。
2.2 烘烤特性最优模型选择与检验
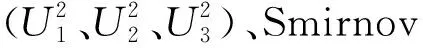
2.2.2 耐烤性遗传模型 通过IECM算法获得烤烟耐烤性RIL群体5类38种遗传模型的AIC值。根据AIC准则,4MG-AI、MX2-AI-AI、MX2-AI-A、MX3-AI-A 4种模型的AIC值较小,可作为备选模型(表2)。依据统计量的适合性检验,发现在P1、P2和RIL的15个检验统计量中,4个模型中有统计学意义的统计量个数分别为5、4、4、4。综合确定耐烤性最优遗传模型为MX2-AI-AI,即2对加性-上位性主基因+加性-上位性多基因模型(表3)。

表2 重组自交系群体烘烤特性指标不同遗传模型的AIC值
续表2 Continued table 2
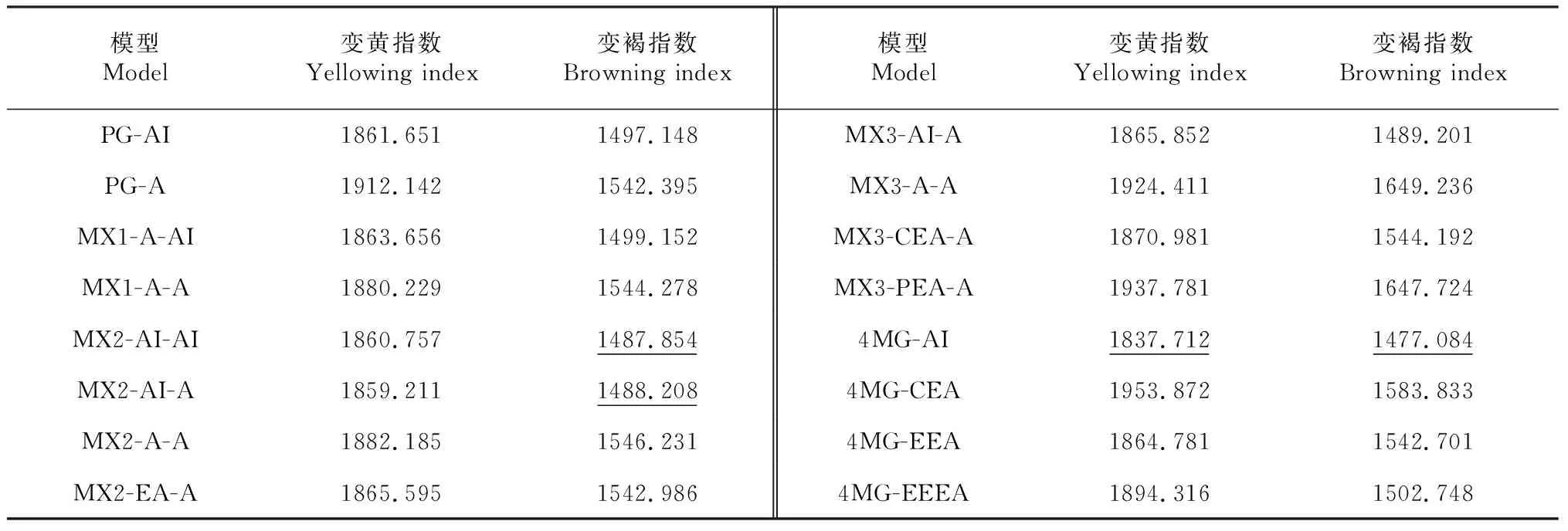
模型Model变黄指数Yellowing index变褐指数Browning index模型Model变黄指数Yellowing index变褐指数Browning indexPG-AI1861.6511497.148MX3-AI-A1865.8521489.201PG-A1912.1421542.395MX3-A-A1924.4111649.236MX1-A-AI1863.6561499.152MX3-CEA-A1870.9811544.192MX1-A-A1880.2291544.278MX3-PEA-A1937.7811647.724MX2-AI-AI1860.7571487.8544MG-AI1837.7121477.084MX2-AI-A1859.2111488.2084MG-CEA1953.8721583.833MX2-A-A1882.1851546.2314MG-EEA1864.7811542.701MX2-EA-A1865.5951542.9864MG-EEEA1894.3161502.748
注:带下划线的数字表示备选模型对应的AIC值。
Note:The numbers with underline refer to the AIC values for the candidate genetic models.MG=major gene model, MX=mixed major gene and polygene model,PG=polygene model, A=additive effect, I=interaction (epistasis), E=equal, C=completely, P=partly.
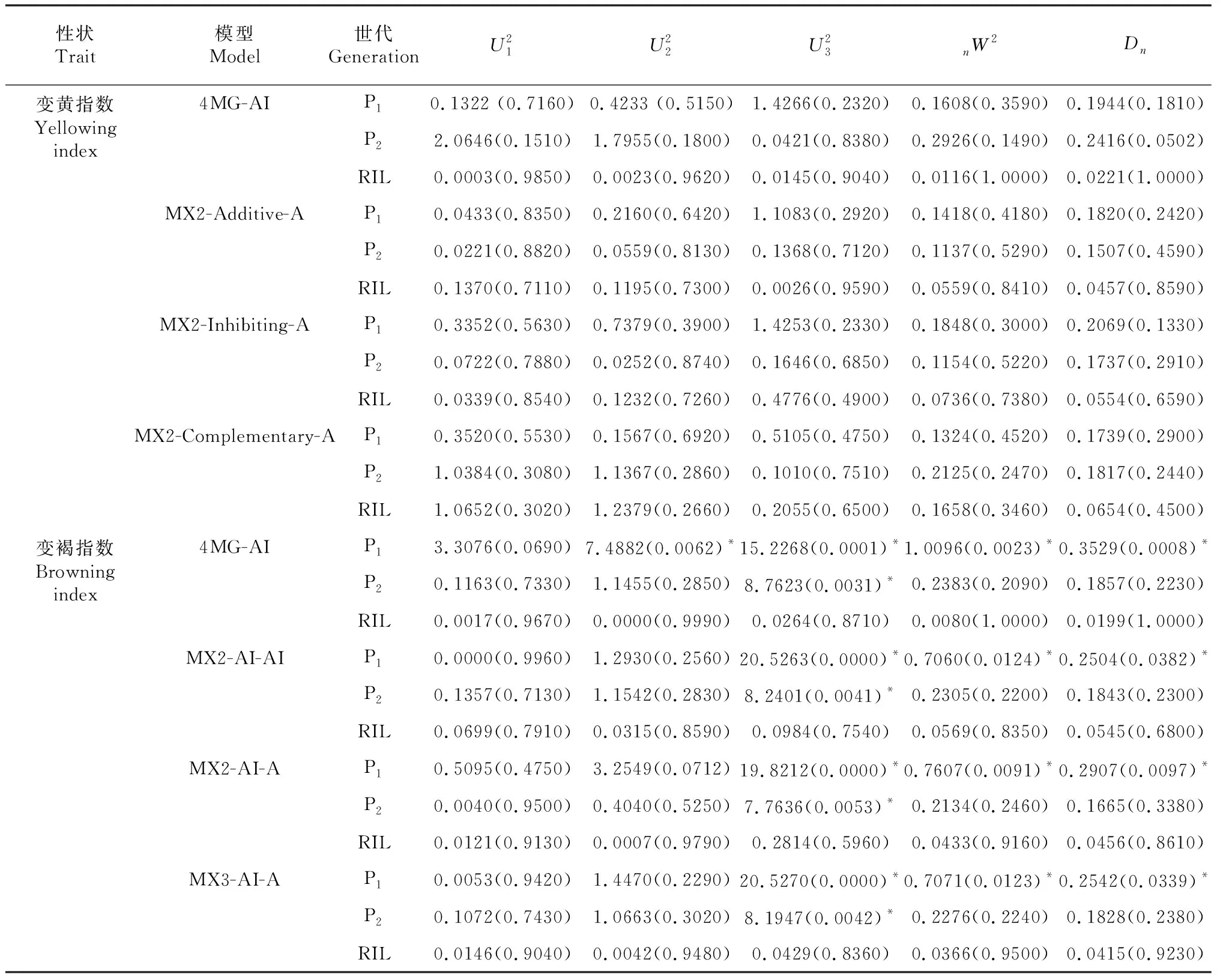
表3 烘烤特性指标备选模型的适合性检验


2.3 烘烤特性的遗传效应分析
2.3.1 易烤性的遗传效应分析 通过对RIL群体变黄指数的分析得出,云烟85×大白筋599组合的易烤性受4对加性-上位性主基因控制,第1、2对主基因的遗传效应较大,分别为12.89、8.80,易烤性增加的效应来源于母本云烟85。第4对主基因的加性效应和基因间的互作效应均为负值,第3、4对主基因间的互作效应较小,其余主基因之间的互作效应明显,主基因遗传率达到95.98 %(表4)。

表4 烘烤特性指标的模型遗传参数

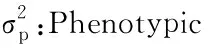
2.3.2 耐烤性的遗传效应分析 对RIL群体变褐指数的遗传分析得出,烤烟组合的耐烤性受2对加性-上位性主基因+加性-上位性多基因控制,2对主基因的加性效应均为负值,不耐烤的效应来源于父本大白筋599,主基因间的互作为正向效应,主基因遗传率为54.26 %,多基因的遗传率为17.82 %,主基因加多基因的遗传率为72.08 %(表4)。
3 讨 论
RIL群体烘烤特性的遗传分析表明易烤性的遗传符合4对加性-上位性主基因模型,遗传效应以第1、2对主基因的加性效应为主,易烤性增加的效应来源于母本云烟85,主基因遗传率很高,有利于QTL位点的发掘和应用。谭效磊[8]通过SSR分子标记技术构建遗传图谱并进行QTL定位,发现4个与易烤性相关的QTL,均为正向加性效应,可解释的表型变异范围为7.57 %~9.26 %。D.Gong等[11]利用F2、F2∶3群体的表型结果,定位到2个共同的易烤性主效QTL,可解释的表型变异率分别为19.3 %~27.3 %、15.2 %~25.1 %,并将其定位在烟草基因组的第6个连锁群上。
RIL群体的耐烤性遗传符合2对加性-上位性主基因+加性-上位性多基因模型,主基因均为负向加性效应,基因间互作效应明显,主基因遗传率达到54.26 %。郝贤伟[10]用联合分离分析方法,分析控制烤烟耐烤性遗传的主基因以负向加性效应为主,主基因之间有明显的互作效应,并利用200个F2群体定位到2个分别位于第3和第9连锁群的耐烤性相关主效QTL,可解释的耐烤性变异率分别为47.01 %和17.49 %,这与RIL群体耐烤性的遗传分析结果基本一致。王春凯等[23]基于全基因组关联分析对烤烟烘烤特性位点进行发掘,并对重要关联区段进行基因筛选和分析,初步预测了4个与易烤性相关的基因和1个与耐烤性相关的基因。
烤烟烘烤特性的遗传分析表明易烤性和耐烤性主要受2对主效基因的控制,这与前人的研究结果[8-11]整体一致,又不尽相同。造成结果差异的原因主要有两方面:一方面数量性状的遗传往往比较复杂,影响烘烤特性的因素较多,所以烘烤特性的遗传也不例外。另一方面,试验群体和模型的不同也会对具体结果造成影响[24]。本研究基于RIL永久群体,采用拓展到4对主基因的数量性状“主基因+多基因”遗传分析方法[25]分析烘烤特性的遗传效应,有利于发掘更多的主基因,提供更多的遗传参数。且RIL群体的不同家系经过重组,基因型得到了固定,这种永久性的分离群体是构建遗传连锁图谱和QTL定位研究的理想材料[26-27]。利用RIL群体进行烤烟烘烤特性的遗传分析,不需考虑基因的显性效应,有利于对控制数量性状基因的加性效应和上位性效应进行估计,为烘烤特性QTL定位提供理论依据。今后,应在已有研究基础上,利用RIL群体和高通量测序技术对烘烤特性QTL进行精细定位,结合最新烟草基因组测序数据,研究将主效QTL定位到相应染色体区段上,进行基因的标记辅助选择和分离克隆。
4 结 论
利用RIL群体对烤烟烘烤特性进行遗传分析,结果表明烤烟易烤性的遗传符合4对加性-上位性主基因模型,主基因遗传率达到95.98 %。耐烤性的遗传符合2对加性上位性主基因+加性上位性多基因模型,主基因之间互作效应明显,主基因遗传率为54.26 %,多基因的遗传率为17.82 %。
猜你喜欢
区域治理(2022年40期)2022-11-27
现代畜牧科技(2021年4期)2021-07-21
湖北农机化(2020年4期)2020-07-24
流行色(2020年9期)2020-07-16
活力(2019年19期)2020-01-06
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
家庭影院技术(2018年9期)2018-11-02
中国烟草学报(2017年1期)2017-11-16
