
不同芹菜品种镉吸收和运转差异研究
2018-10-17 09:19徐瑞深乜兰春王珊珊王若男
植物营养与肥料学报 2018年5期
徐瑞深,乜兰春,王珊珊,李 潘,王 政,王若男
(河北农业大学园艺学院/河北省蔬菜种质创新与利用重点实验室/河北省蔬菜产业协同创新中心,河北保定 071001)
土壤重金属污染已经成为影响蔬菜产品品质安全的重要因素之一。据统计,我国受重金属污染的耕地面积已超过2000 万hm2,约占耕地总面积的1/5,镉 (Cd) 是环境中毒性最强、对菜地污染最为严重的重金属之一[1–2],可通过有机肥、灌溉水、大气沉降、农业和动物废物以及农药等多种途径进入到土壤中[3]。Cd具有化学活性强、移动性大、生物毒性强且持久的特性,易被植物吸收,能在植物体内残留,对植物产生毒害,并通过食物链的富集危及人类健康[2,4–6]。
由于植物的遗传特异性,不同种类以及同一种类不同品种蔬菜之间对于重金属Cd的吸收和积累存在显著差异,叶菜类蔬菜较其他种类蔬菜更易富集Cd[7]。芹菜是深受人们喜爱的一种叶菜类蔬菜,在我国广泛种植。此前已有研究报道,芹菜对重金属Cd具有相对较强的富集能力[6–7]。但关于芹菜Cd积累的品种差异研究报道并不多。开展镉积累品种差异及机理研究是筛选耐镉且食用器官低积累型品种、开展相关育种及生产调控、减少镉吸收与积累、保证产品质量安全的基础。Cd进入植物体首先通过根系,根部离子流速与植物体内的离子积累量之间存在着必然的联系[8]。非损伤微测技术 (noninvasive micro-test technology,NMT) 能够在正常生理状态下快速准确地检测离子流速的变化,直观地反映植物对离子的吸收情况。本文在研究Cd胁迫下不同品种芹菜生长、Cd积累及转运差异的基础上,利用非损伤微测技术分别对高、低Cd积累型的两个品种芹菜根部Cd2+流速进行实时检测。以期为筛选低积累型芹菜品种和揭示芹菜Cd积累品种差异的内在生理机制奠定基础。
1 材料与方法
1.1 试验设计
Cd胁迫下芹菜生长及Cd积累的品种差异研究采用盆栽试验,于2016年3—6月在河北农业大学日光温室内进行,供试的10个芹菜品种情况及种子来源见表1。播种前,将各品种种子用清水浸种12 h,再于20℃恒温培养箱中催芽7 d,播于穴盘。育苗14 d后,挑选长势一致的芹菜幼苗移栽至装有蛭石的花盆 (直径25 cm × 高20 cm,实际装入蛭石体积为6.5 L) 中,每盆10株,每品种6盆,其中3盆浇灌含15 mg/L氯化镉 (CdCl2) 的1/2华南农业大学叶菜类营养液,配方为:Ca(NO3)2·4H2O 472 mg/L、KNO3202 mg/L、NH4NO380 mg/L、KH2PO4100 mg/L、K2SO4174 mg/L、MgSO4·7H2O 246mg/L、EDTA-NaFe 36.6 mg/L、H3BO42.86 mg/L、MnSO4·4H2O 2.13 mg/L、ZnSO4·7H2O 0.22 mg/L、CuSO4·5H2O 0.08 mg/L、(NH4)6Mo7O24·4H2O 0.02 mg/L),进行Cd胁迫处理;另3盆浇灌等量1/2华南农业大学叶菜类营养液作对照,每3 d浇灌一次,每次浇灌0.2 L,累计浇灌15次,每盆Cd施用量为45 mg,按盆内装与蛭石等量的菜园土计算,土壤Cd含量2.7 mg/kg,模拟超过《土壤环境质量标准》(GB15618-1995) 三级标准,达到重污染水平的土壤。移栽至花盆45 d后,将芹菜植株从基质中取出,自来水冲洗干净后,再用超纯水浸泡三次,每次10 min,吸水纸吸干水分,分别测定基部短缩茎茎粗和单株叶柄数、叶柄基部粗度和单株复叶数。之后,将根、叶柄和叶片分别装入信封中,在105℃杀青30 min,65℃下烘干至恒重,称重,用于测定Cd含量。计算相对生长量 (relative growth yield,RGY)和转运系数 (translocation factor,TF)[9]。
相对生长量 (RGY) = 处理生长指标/对照生长指标
转运系数 (TF) = Cd胁迫处理下植株地上部Cd含量/胁迫处理植株根系Cd含量
根部Cd2+离子流品种差异研究分别以筛选出的Cd积累量较低的‘速生香芹’和Cd积累量较高的‘种都金黄芹菜’为试材,种子处理同上,穴盘基育苗14 d后,选用长势一致的幼苗移栽至水培盆中。采用华南农业大学叶菜类营养液配方进行营养液栽培,21 d后分别测定根系分生区、伸长区和根毛区Cd2+离子流速。
1.2 测定项目与方法
1.2.1 Cd含量测定 取烘干的芹菜叶片、叶柄和根系样品1.0 g进行微波消解,消化剂为25 mL HNO3:HClO4(5∶1),逐步升温,依次为:110℃,5 min;130℃,5 min;150℃,5 min;280℃,约20 min。280℃消煮液体颜色需显示无色透亮,冷却后过滤,超纯水定容至50 mL容量瓶。将上述植物消煮液采用植物全量–等离子体原子发射光谱 (ICP-OES) 法测定Cd含量,重复3次。
1.2.2 根系Cd2+离子流速测定 Cd2+离子流速采用非损伤微测系统[non-invasive micro-test system (Younger USA LLC, Amherst, MA 01002, USA)]测定。采用Cd2+选择性微传感器获取进出芹菜根系测定点的Cd2+离子浓度 (mmol/L级)、流速和流向信息。测试前将测试样品放置于测试液中平衡10 min,测试液配方为 0.1 mmol/L CaCl2、0.1 mmol/L KCl、0.05 mmol/L CdCl2、0.3 mmol/L MES、pH 5.8。测试点如图1所示:分生区为静止中心向下100 μm,伸长区为静止中心向下450 μm,成熟区为第一根根毛下移400 μm的主根处。每个处理重复6次,每次测定10 min。
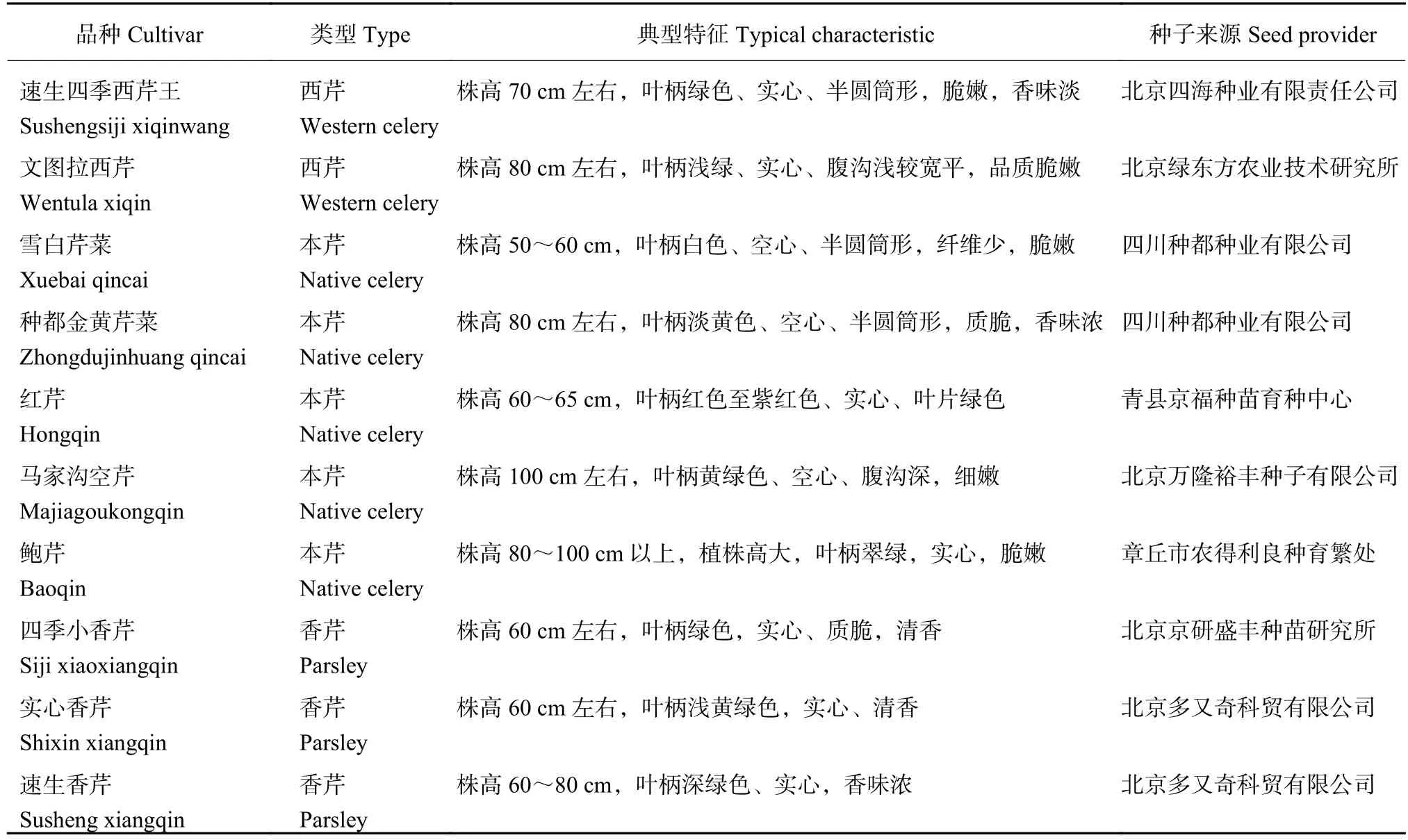
表1 试验材料Table 1 Experimental materials
1.3 数据处理
采用Microsoft Excel 2016对试验数据进行统计整理,采用DPS 7.05 软件对数据进行方差分析和显著性测验。
2 结果与分析
2.1 Cd胁迫对不同品种芹菜生长的影响
Cd胁迫下,芹菜生长受到抑制,普遍表现为植株细弱、变矮、叶片发黄等。由表2可以看出,在Cd胁迫下,‘红芹’ 、‘鲍芹’和‘种都金黄芹菜’地下部干重与对照差异不显著;其余品种地下部干重均显著低于对照,相对生长量RGY在0.33~0.78,其中‘速生四季西芹王’ 、‘文图拉西芹’和‘四季小香芹’最低,分别只有0.33、0.36和0.37。
从地上部生长指标来看,Cd胁迫下,‘四季小香芹’株高高于对照,‘速生四季西芹王’和‘雪白芹菜’与对照差异不显著,其余品种均显著低于对照,RGY在0.76~0.93,‘种都金黄芹菜’最低。‘四季小香芹’短缩茎粗显著高于对照,其它品种显著低于对照,RGY在0.45~0.93,以‘种都金黄芹菜’最低,其次为‘雪白芹菜’和‘鲍芹’ 。所有品种的平均叶柄粗均显著低于对照,RGY在0.44~0.85,也以‘种都金黄芹菜’最低。‘四季小香芹’和‘文图拉芹菜’叶柄数多于对照,‘种都金黄芹菜’少于对照,其余品种与对照差异不显著。复叶数除‘马家沟空芹’的少于对照外,其余品种与对照差异均未达显著水平。可见,Cd对芹菜地上部生长的抑制作用主要表现在使短缩茎及叶柄变细、株高变矮,对叶柄数和叶片数影响较小。
从地上部干重来看,Cd胁迫下‘四季小香芹’地上部干重显著高于对照,RGY达1.87,‘速生四季西芹王’ 、‘文图拉芹菜’ 、‘实心香芹’和‘雪白芹菜’与对照差异不显著。而‘种都金黄芹菜’ 、‘红芹’ 、‘鲍芹’ 、‘马家沟芹菜’和‘速生香芹’均显著低于对照,RGY在0.57~0.81,其中‘种都金黄芹菜’最低。可见,不同品种芹菜地上部和地下部对Cd胁迫响应存在显著差异。
综合以上结果,在本试验条件下,镉胁迫使‘速生四季西芹王’、 ‘文图拉芹菜’和‘四季小香芹’地下部受到显著抑制,而地上部生长不受影响,甚至对‘四季小香芹’地上部还有促进作用;而‘种都金黄芹菜’ 、‘红芹’ 、‘鲍芹’地上部生长受到显著抑制,地下部则无显著影响;‘马家沟芹菜’和‘速生香芹’地上部和地下部生长均受到抑制。考虑芹菜产量由地上部构成,‘四季小香芹’ 、‘速生四季西芹王’ 、‘文图拉芹菜’ 、‘实心香芹’和‘雪白芹菜’为耐镉型;而‘种都金黄芹菜’ 、‘红芹’ 、‘鲍芹’ 、‘马家沟空芹’和‘速生香芹’为镉敏感型。
2.2 Cd胁迫下不同品种芹菜Cd积累的差异
从表3可以看出,10个品种的对照叶片、叶柄和根中Cd含量很低,在1.0~2.0 mg/kg之间。而在Cd胁迫下,各品种叶片、叶柄和根系中Cd含量增加10倍左右,且不同品种间差异显著。叶片Cd含量以本芹‘种都金黄芹菜’和‘雪白芹菜’最高;而香芹中的‘实心香芹’和‘速生香芹’叶片Cd含量最低,分别比‘种都金黄芹菜’低32.31%、31.05%。叶柄Cd含量也以‘种都金黄芹菜’最高,其次为‘雪白芹菜’ ;‘速生香芹’叶柄Cd含量最低,比‘种都金黄芹菜’低54.66%。根系Cd含量以‘四季小香芹’和‘红芹’最高,‘鲍芹’和‘雪白芹菜’最低,分别比‘四季小香芹’低51.96%、49.96%。芹菜以叶柄为食用器官,无论从单位干重叶柄Cd含量看,还是从单株叶柄及全株Cd积累量 (表4) 看,‘雪白芹菜’和‘种都金黄芹菜’为高Cd积累型;‘速生香芹’和‘实心香芹’为低Cd积累型。

表2 Cd 处理对 不同 品种 芹菜生 长的 影响Table 2 Effects of the Cd treatments on the growth of different cultivars of celery
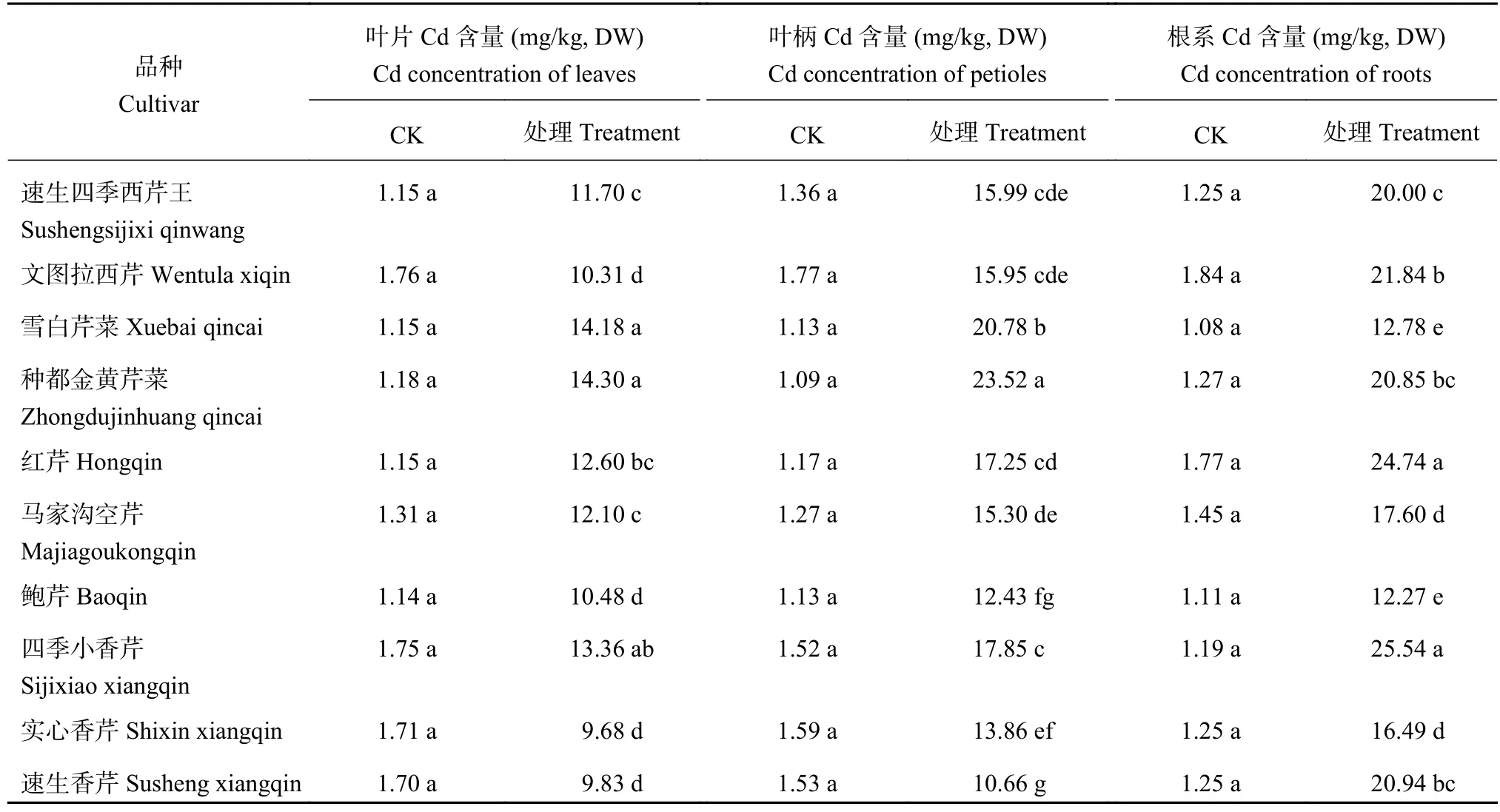
表3 芹菜不同部位重金属Cd含量Table 3 Cd concentrations in different parts of celery
2.3 Cd胁迫下不同品种芹菜Cd转运系数的差异
从图2可以看出,Cd转运系数以‘雪白芹菜’最高,达1.39,显著高于其他品种,其次为‘种都金黄芹菜’和‘鲍芹’ ,分别为0.96和0.94。‘速生香芹’转运系数最低,仅为0.48,比‘雪白芹菜’低65.47%,‘四季小香芹’ 、‘红芹’ 、‘文图拉芹菜’转运系数也较低。可见,芹菜不同品种,由根系向地上部的Cd转运能力存在显著差异。地上部Cd含量高的‘种都金黄芹菜’和‘雪白芹菜’对Cd转运能力也最高;而地上部Cd含量低的‘速生香芹’对Cd转运能力也最低。
2.4 Cd胁迫下高、低积累型芹菜根部Cd2+ 离子流的差异
Cd2+流速为负值表示为内流即吸收。由图3可以看出,‘速生香芹’根部Cd2+离子流速分生区 >伸长区 > 成熟区,其中分生区和伸长区随着时间延长有逐渐减小的趋势。而成熟区的Cd2+离子流速相对稳定,在15~30 pmol/(cm2·s) 的范围内。‘种都金黄芹菜’根部分生区和伸长区的Cd2+离子流速也随测试时间逐渐减小,而成熟区的Cd2+离子流速则有缓慢增加的趋势,最高达到60.26 pmol/(cm2·s)。成熟区是根系吸收水分和矿质元素的主要部位,‘种都金黄芹菜’根部成熟区Cd2+离子流速高,说明‘种都金黄芹菜’根部具有较强的Cd吸收能力,这与其地上部Cd含量及积累量高密切相关。
3 讨论与结论
大量研究表明,不同品种作物在重金属Cd胁迫下会产生不同的响应[10-12]。蔬菜上,小白菜[13,14]、空心菜[15]、大葱[16]、番茄[17-18]和苋菜[19-20]等均有相关报道。Yang等[21]和肖旭峰等[22]研究表明,随着Cd处理浓度增加,Cd对芹菜生长产生的抑制作用会增强。Zhang等[23]报道,Cd胁迫下不同品种芹菜地上部生长和Cd积累量产生不同的响应,27个品种中有20个品种地上部生物量受到抑制,另外7个品种与对照无显著差异甚至有促进作用。本研究也表明,芹菜耐Cd性存在品种差异,Cd胁迫下,‘四季小香芹’地上部生长量显著高于对照,‘速生四季西芹王’ 、 ‘文图拉芹菜’ 、 ‘实心香芹’和‘雪白芹菜’地上部生长量与对照差异不显著,这5个品种为耐镉型;而‘种都金黄芹菜’ 、 ‘红芹’ 、 ‘鲍芹’ 、‘马家沟芹菜’和‘速生香芹’地上部生长量受到显著抑制,为镉敏感型。本研究还发现,同一品种的地上部和地下部对Cd胁迫的响应也存在显著差异。镉胁迫下,‘速生四季西芹王’ 、‘文图拉芹菜’和‘四季小香芹’地下部受到显著抑制,而地上部生长不受影响,甚至对‘四季小香芹’地上部还有促进作用;而‘种都金黄芹菜’ 、‘红芹’ 、‘鲍芹’地上部受到显著抑制,对地下部则无显著影响。这可能与Cd转运及不同器官耐Cd性不同有关,Cd由地下部向地上部转运得多,地上部就有可能受影响;反之,向地上部转运少,Cd主要集中在根部,根系发育就有可能受到影响,而地上部生长受影响小。不同器官耐镉性也可能存在差异。植物器官耐重金属机理包括螯合重金属[24]、促进重金属的排出[25]、区室化重金属[26]、诱导相关酶活性[27]及基因表达等,关于芹菜不同品种及不同器官耐镉机理有待进一步研究。
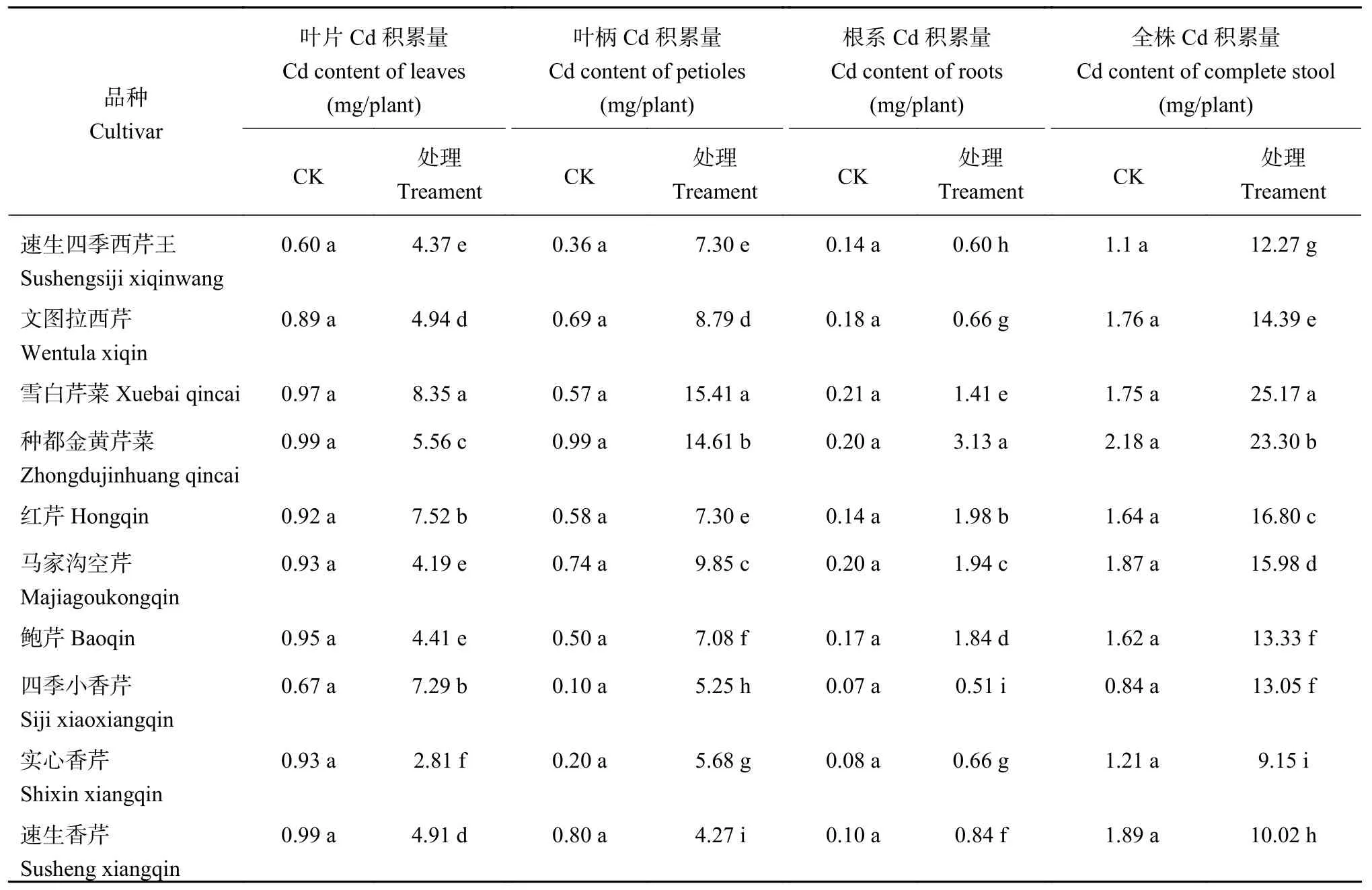
表4 芹菜不同部位重金属Cd积累量Table 4 Cd accumulation amounts in different parts of celery
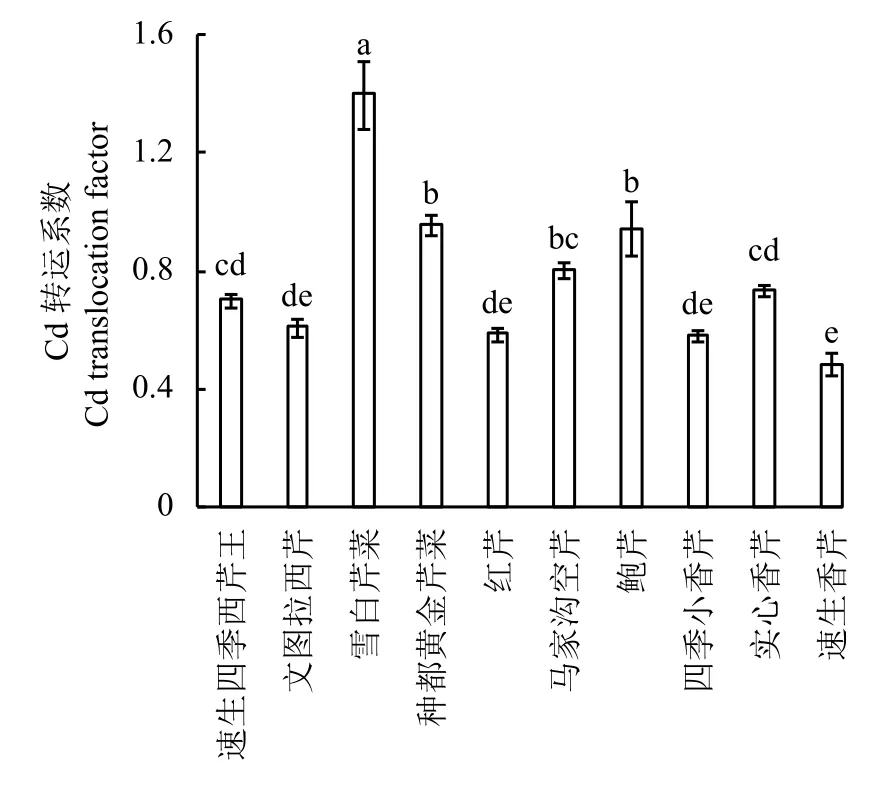
图2 不同品种芹菜Cd转运系数Fig. 2 Translocation factors of different cultivars of celery

图3 Cd胁迫下‘速生香芹’和‘种都金黄芹菜’根系不同部位Cd2+ 流速变化Fig. 3 Cd2+ flux kinetics in different regions of roots of ‘Susheng xiangqin’ and ‘Zhongdujinhuang qincai’
不同品种甘薯可食用部位Cd积累差异可高达87倍[28]。而萝卜不同品种间可食用部位Cd积累差异很小[29]。陈瑛等[14]从60个小白菜品种中筛选出12个植株体内Cd含量低、生物量受Cd影响小的小白菜品种。本研究表明,芹菜食用器官叶柄中Cd含量以‘种都金黄芹菜’和‘雪白芹菜’最高,‘速生香芹’和‘实心香芹’最低。结合不同品种对Cd胁迫的生长响应可以看出,‘雪白芹菜’表现为高Cd含量和高生长量,‘种都金黄芹菜’表现为高Cd含量和低生长量,‘速生香芹’表现为低Cd含量和低生长量,‘实心香芹’表现为低Cd含量和高生长量。这种生长响应与Cd含量关系的品种差异可能与不同品种芹菜耐Cd和解除Cd毒害的能力不同有关,但其分子机理还有待进一步研究。本试验中‘实心香芹’作为耐镉且食用器官低积累型品种在生产和耐镉芹菜种质创新中具有较高的应用价值。
植物体内Cd含量取决于根系吸收能力和体内转运能力[30-32]。根系Cd吸收能力与木质部的转运能力共同对苋菜的Cd积累产生影响[20];Wang等[33]研究表明,相对于Zn和Ni,Cd在芹菜根系具有更强的转运能力。Zhang等[23]研究表明,不同品种芹菜具有不同的转运能力,可食用部位Cd含量较高的芹菜品种具有更强的Cd转运能力。本研究也证实,地上部Cd含量高的‘种都金黄芹菜’和‘雪白芹菜’根对Cd转运能力也最高;而地上部Cd含量低的‘速生香芹’对Cd转运能力也最低。Cd进入植物体首先通过根系,根部离子流速与植物体内的离子积累量之间存在着必然的联系[8],但目前关于芹菜根系对Cd2+离子吸收能力的品种差异还未见报道。本研究利用非损伤微测技术 (NMT) 分别对高、低Cd积累型的两个品种芹菜根部Cd2+流速进行实时检测,结果表明,高Cd积累型的芹菜根系成熟区 (根毛区) Cd2+离子流速高于低Cd积累型的芹菜,说明高积累型芹菜品种根部具有更强的Cd吸收能力。Cd积累的品种差异是筛选低积累品种的主要依据。传统的研究方法是对植株进行Cd胁迫,再分析Cd积累量的差异,由于试验周期长,易受不可控因素的干扰,降低了低积累品种的选育效率,Cd2+流速直接反映了植物的主动吸收能力,可以考虑以Cd2+流速为依据,从Cd2+流速慢的品种中更容易筛选出地上部Cd含量低的品种或材料,从而提高筛选效率。
猜你喜欢
红蜻蜓·高年级(2022年10期)2022-12-31
中医药临床杂志(2022年2期)2022-11-15
小天使·四年级语数英综合(2022年6期)2022-06-21
农业灾害研究(2022年2期)2022-05-31
南方农业·下旬(2022年4期)2022-05-24
作文·小学低年级(2021年4期)2021-11-02
作文小学中年级(2021年4期)2021-05-07
林业与生态(2019年6期)2019-07-02
微型小说选刊(2016年6期)2016-12-08
食品工业科技(2014年23期)2014-03-11
