
二穗短柄草Trihelix转录因子家族的基因组学分析
2018-10-16 07:13:34胡风越刘廷武周曼丽周颖君杨立明纪剑辉
江苏农业科学 2018年18期
胡风越,刘廷武,周曼丽,周颖君,周 芮,杨立明,纪剑辉
(淮阴师范学院生命科学学院,江苏淮安223300)
在植物的转录因子家族中,研究得较为全面的是与逆境相关的转录因子,如 ABF、MYB、NAC、DREB、DOF、WRKY 等转录因子家族,对这些转录因子相关基因的恰当的转化运用,可以提高植物对逆境胁迫的耐受性[1-6]。Trihelix转录因子是与抗逆相关的转录因子家族之一,目前在拟南芥、水稻、棉花、高粱等植物中均有相关研究。研究显示,拟南芥中有30个Trihelix转录因子,而水稻中含有31个[7]。此外,由于Trihelix转录因子在光应答反应、植物生长发育和逆境胁迫中显著的调控作用[8-12],它们对生产实践中的品种改良应用具有重要意义。二穗短柄草由于具有植株矮小、生育期短、遗传转化率高以及其与小麦基因组相似度高达95%、能同样感染小麦易感的各种细菌的特点,成为继拟南芥和水稻之后新型模式植物的理想植株,并能有效弥补模式植物水稻的不足[13-14]。本研究利用生物信息学分析手段,对二穗短柄草Trihelix转录因子家族基因进行结构特征、进化情况及染色体定位等分析,并对该基因家族在水稻、二穗短柄草以及玉米等物种染色体区段的复制作了分析,以期更全面地了解禾本科植物Trihelix基因家族。1 材料与方法
1.1 数据的收集与分析
在植物转录因子数据库(Plant Transcription Factor Database,简称TFDB)中获取水稻、二穗短柄草、玉米、高粱及拟南芥中的Trihelix基因序列,随后对相关数据进行整合分析,利用Pfam方法在水稻中检测是否存在此家族蛋白的特征结构域,借助同样的方法,完成对Trihelix特殊结构域的序列筛选,进而得到不同物种Trihelix蛋白的全部序列[15]。利用ExPASy数据库(http://www.expasy.org/)对二穗短柄草Trihelix蛋白的分子量、染色体位置、等电点等基因组信息进行在线分析[16]。
1.2 保守基序(motif)的鉴定与分析
根据 Trihelix家族保守结构域特点,在 MEME程序(http://meme.nbcr.net/meme/cgi- bin/meme.cgi)中,将参数设置成“any”,设定这些基序的长度为10~200,预期有20个基序[17],来分析其蛋白质保守基序。
1.3 Trihelix转录因子家族系统进化树的构建
首先对Trihelix家族特殊保守结构域的氨基酸进行多序列联配分析,应用Cluster 3.0软件分析所得结果,随后借助Mega软件(http://www.megasoftware.net/)对二穗短柄草Trihelix保守结构域作进一步分析,Bootstrap的值设定成1 000[18],随后将 Mega 程序分析的结果借助 WebLogo 3.3 软件进行图示化,即可验证转录因子保守域的分析结果。依据二穗短柄草Trihelix特殊结构域分析的方法和步骤,采用邻接法构建出水稻、拟南芥、玉米、二穗短柄草和高粱中该家族的系统进化树[19],发现在不同物种中,Trihelix家族均主要分为5个亚类。这一结果显示,在植物不同物种中,Trihelix转录因子家族基因仍具有较高的保守性。
1.4 Trihelix基因家族的染色体区段复制分析
利用植物基因组复制数据库(PGDD,网址为 http://chibba.agtec.uga.edu/duplication/)对所选的 Trihelix 基因各自所在染色体区段的线性复制情况[20]进行分析,将所有存在染色体复制关系的数据进行整理,借助基因染色体定位软件MapInspect完成图示化;此外,按照上述方法将二穗短柄草与水稻、二穗短柄草与高粱、二穗短柄草与玉米的Trihelix基因利用植物基因组复制数据库进行线性复制关系分析,相关结果借助Circos软件进行图示化[21],便可直观地看出所需检测的染色体之间的同源性关系。
2 结果与分析
2.1 二穗短柄草Trihelix转录因子家族的鉴定
在二穗短柄草基因组数据库中共鉴定出28个Trihelix转录因子。对其蛋白质大小、分子量、等电点以及染色体位置等基本信息进行分析,结果显示其中氨基酸序列最长的有975个,最短的有128个,分子量为 14.01~96.63 ku,等电点为3.976 3~11.578 9。蛋白结构域保守型分析结果显示,氨基酸残基色氨酸(Trp,简称 W)-2、W -63、半胱氨酸(Cys,简称C)-99(2、63、99分别表示在此氨基酸序列上的第2、63、99位)具有极高保守性(图1),且短柄草所有Trihelix转录蛋白中均包含典型的螺旋-环-螺旋保守结构。
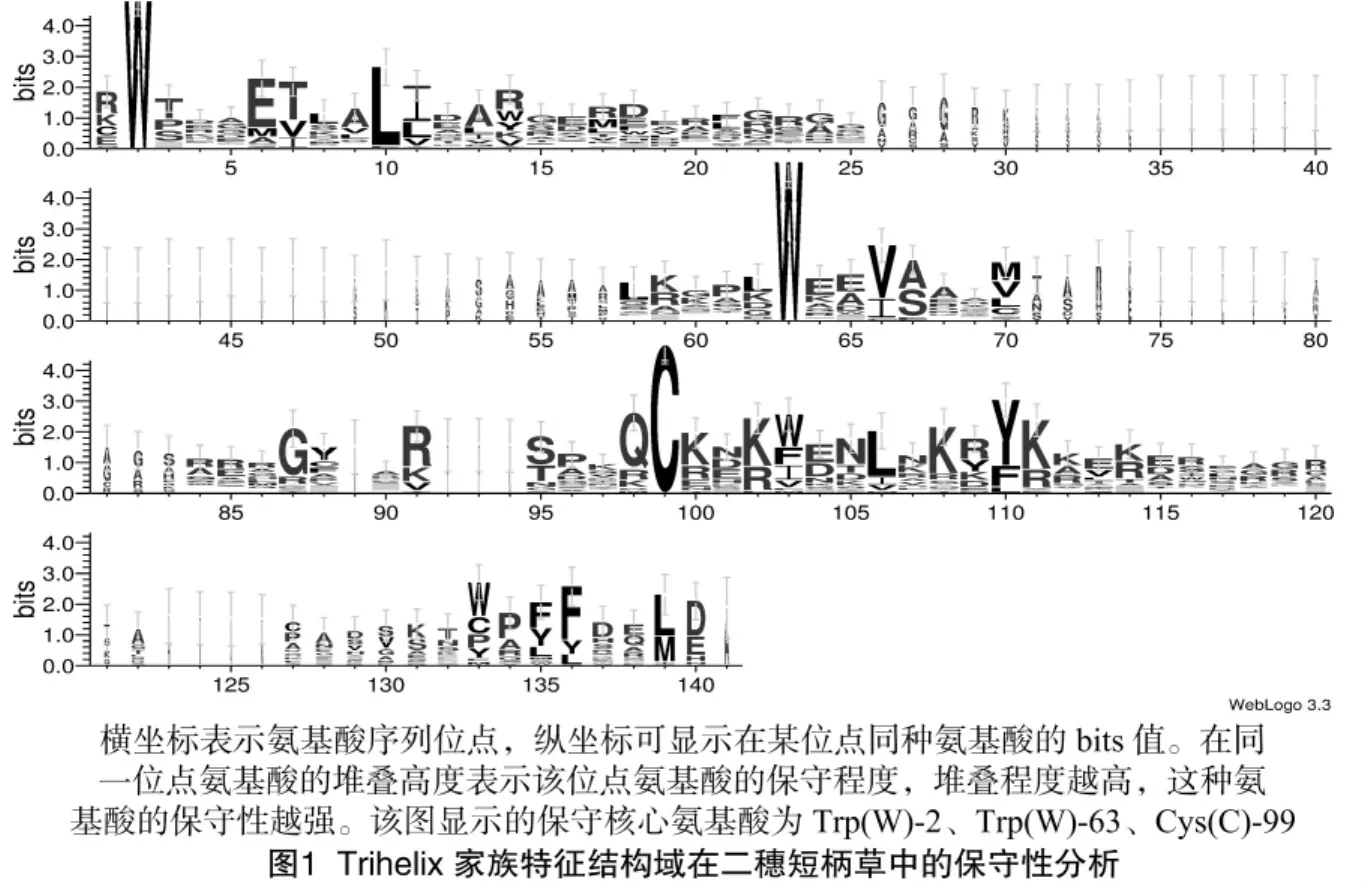
2.2 二穗短柄草Trihelix转录因子家族的进化分析
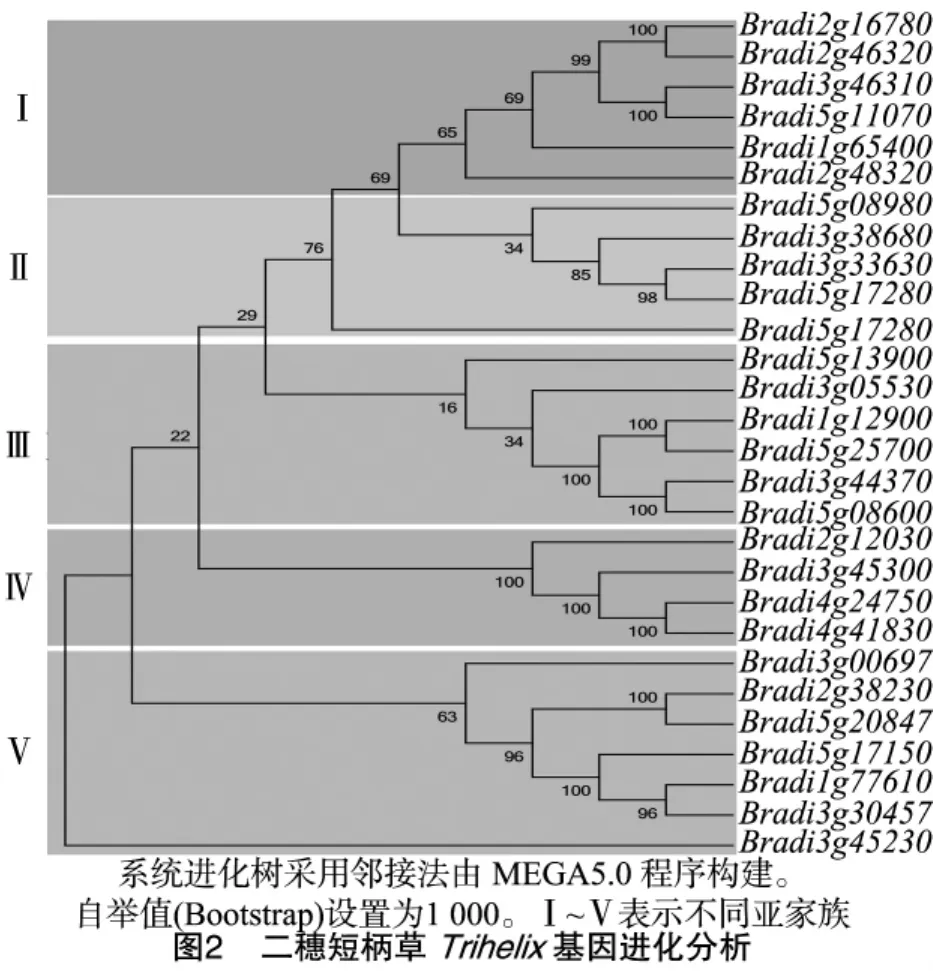
依据Trihelix蛋白保守序列特点,在MEME程序中采用邻接法构建了二穗短柄草Trihelix基因家族的进化树(图2),结果将二穗短柄草Trihelix基因家族分为5个亚类。在数量分布上,亚家族Ⅳ中含有最少的Trihelix转录因子,而亚家族Ⅴ含有的Trihelix转录因子最多;在分支结构上,亚家族Ⅰ和Ⅱ的同源关系相对较近,且可看出在亚家族Ⅴ上的Bradi3g45230基因与其他Trihelix基因的亲缘关系较远。此外,对水稻、拟南芥、二穗短柄草和高粱中的Trihelix蛋白进行聚类分析发现,Trihelix家族均分为5个亚类(Ⅰ~Ⅴ)(图3),且每个亚类中均有Trihelix转录因子分布,由此可清晰地看出各物种中Trihelix转录因子之间的同源关系。高粱、玉米的Trihelix基因进化距离较长,分别有近71%、69%的基因在进化树的末端才分支,因而推测这2个物种Trihelix基因的进化较晚,其次是二穗短柄草和拟南芥,最早的是水稻。
2.3 二穗短柄草Trihelix家族全长蛋白的结构性分析
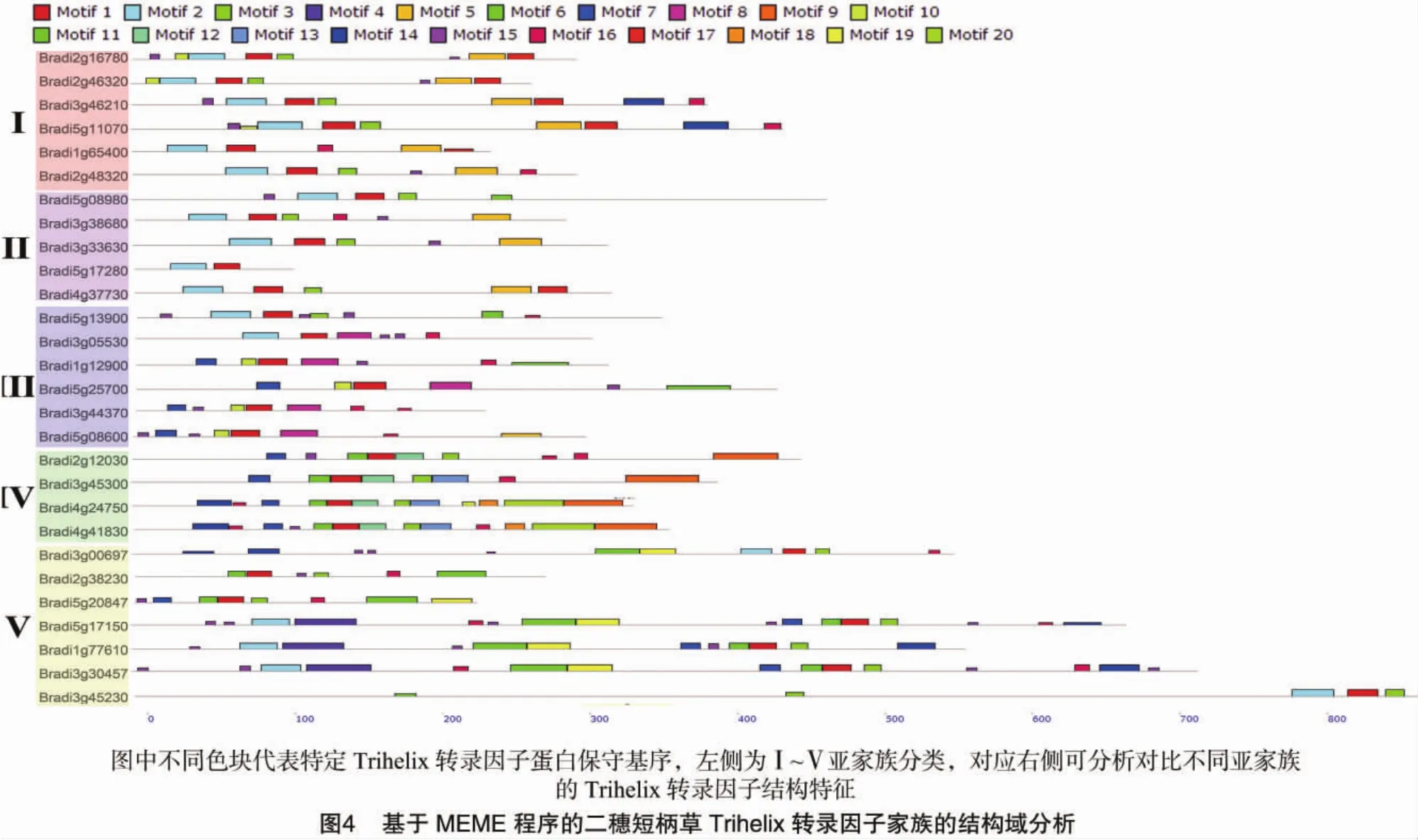
基于MEME程序的蛋白结构域分析结果显示,二穗短柄草的28个Trihelix转录因子同样可被分为5个亚家族,与上述进化树分析结果一致。就同一亚家族内结构特点分析可知:亚家族Ⅰ中都含有保守基序Motif2、Motif1和Motif5,亲缘关系更为接近的Bradi3g46210和Bradi5g11070基因表达蛋白的C端都含有Motif16;亚家族Ⅱ中共有的保守基序与亚家族Ⅰ相比不含有Motif5;亚家族Ⅲ中均存在Motif1和Motif8;亚家族Ⅳ的显著特征是含有1段由Motif3、Motif1和Motif12相邻构成的保守序列;亚家族Ⅴ中亲缘关系较近的Bradi3g00697、Bradi5g17150、Bradi1g77610 和 Bradi3g30457均含有Motif 3,均与Motif19紧密相邻,且后3种Trihelix转录因子的同源性更高。在不同亚家族之间,根据Motif5和Motif2判断亚家族Ⅰ和亚家族Ⅱ在系统发生上亲缘关系更接近。此外,在所有Trihelix转录因子中Motif1蛋白序列高度保守(图4)。
2.4 二穗短柄草Trihelix转录因子家族在染色体上的定位
用Mapinspect软件分析图示,可见二穗短柄草的28个保守Trihelix基因在5条染色体上均有分布(图5),在3号染色体上分布最多,有9个Trihelix基因,而在1号和4号染色体上最少,仅有3个。并且,Trihelix基因在同一染色体上还存在聚集现象。如在3号染色体上9个Trihelix基因聚集在3个区段,分别位于0.27、32、46 Mb附近;而5号染色体上所有Trihelix基因均聚集在11~28 Mb区段,即位于染色体长臂区段。
2.5 二穗短柄草Trihelix转录因子在二穗短柄草、高粱、玉米和水稻中的染色体复制分析


由于高粱、水稻和玉米均为单子叶植物,其相互之间亲缘较近,所以通过比较二穗短柄草与这三者之间的染色体复制情况,可以分析Trihelix基因在种内和种间的进化轨迹。复制分析结果见图6,可见二穗短柄草的染色体有5对,首先,其自身同一亚家族的Trihelix基因会形成复制关系,其次,二穗短柄草与高粱(10对染色体)、水稻(12对染色体)和玉米(10对染色体)也形成了线性复制。据此可推测,在物种进化过程中可能发生了染色体加倍、易位,以及基因的复制、重排、缺失和一些同源基因的丢失等进化事件。
3 讨论与结论
Trihelix转录因子又称为GT因子家族[22-23],因其含有3个螺旋环的特殊保守结构而得名,并且其结构与MYB转录因子家族极为相似,但因连接helix的结构不同(“环”或“转角”)而造成功能上的差异,在系统发生上,Trihelix转录因子可能来源于1 个 MYB -like基因[24-25]。
Trihelix转录因子最初只在植物中发现,后来有研究推测可能在人类和果蝇中也存在,陆婷婷研究表明,Trihelix转录因子在动物和植物中有不同的结构,绝大多数植物Trihelix蛋白为单Trihelix功能域蛋白,而动物中单Trihelix功能域占少数,取而代之出现了3个Trihelix功能域蛋白和6个Trihelix功能域蛋白,且多伴有其他功能域如SCAN功能域[由C2H2锌指蛋白家族成员SRE-ZBP、Ctfin51、AW-1(ZNF174)和Number 18组成],以辅助调节动物的生长代谢过程[26]。而在功能上,植物中Trihelix转录因子主导光应答、植物生长发育中的蛋白表达调控以及与抗干旱、耐盐、冷害、病害等逆境胁迫应答相关[11,27-28],动物中 Trihelix蛋白辅助调控生长代谢,具体功能仍待探索。

本研究通过对二穗短柄草Trihelix转录因子的生物信息学分析,共鉴定出5个亚家族共28个Trihelix转录因子基因,水稻中是5个亚类31个Trihelix转录因子基因,二穗短柄草中Trihelix转录因子数量虽不及水稻,但从聚类分析结果来比较,仍具有较高的相似性。水稻、高粱和玉米Trihelix多基因进化关系构建结果同时表明,Trihelix在物种间的进化晚于在物种内的进化;此外,依据Trihelix转录因子在系统进化树上的进化距离,推测Trihelix在高粱和玉米中的进化最晚,在二穗短柄草和拟南芥中次之,最早是在水稻中,但此结论需要进一步研究证实。
综上,通过对单子叶禾本科模式植物二穗短柄草的生物信息学研究,分析其Trihelix转录因子家族基因的鉴定、结构特征、进化特点、染色体定位及复制情况,从而对禾本科植物的Trihelix转录因子有更全面的了解,为小麦等其他未测序物种的基因功能分析奠定了基础。
猜你喜欢
核农学报(2020年6期)2020-06-30 06:15:38
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
科学之谜(2019年3期)2019-03-28 10:29:44
科学之谜(2018年8期)2018-09-29 11:06:46
植物研究(2018年4期)2018-07-24 01:06:24
恋爱婚姻家庭·养生版(2016年9期)2016-09-07 11:25:01
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
中央民族大学学报(自然科学版)(2015年2期)2015-06-09 08:45:16
现代检验医学杂志(2015年2期)2015-02-06 02:01:01
