
4种榕属植物种子生物学特征及幼苗生长发育特性研究
2018-10-15 03:17,,,,,,
种子 2018年9期
, , , , , ,
(1.西南林业大学生物多样性保护与利用学院, 云南 昆明 650224; 2.西南林业大学林学院, 云南 昆明 650224)
榕树是桑科(Moraceae)榕属(Ficus)中的乔木树种的总称[1],为常绿乔木或灌木。全世界的榕树约有750种[2-3],榕属植物主要分布在热带,亚热带地区,它是热带雨林中最大的木本属种之一[4],在滇南热带雨林中就有40多种[5]。榕树是所有物种中结实量最高的,在热带地区,一年四季均会结果,一些种年结实3~8次,是任何一种植物都难以比拟的,在其他植物挂果量稀少的冬春季,它为多种兽类、鸟类、蝙蝠、昆虫、土壤动物和微生物等提供食物和栖息场所,是国际上公认的热带森林生态系统中的一类关键植物[5-6]。除此之外,榕树还与居民的生活密切相关,在滇南佛教文化盛行地区,部分榕树常被当做神树,并对其严加保护,具有重要的宗教文化意义[5]。同时榕树在食用、药用和工业上也具有开发价值,是重要的资源植物[7]。榕树是我国南方城市常用的行道树种和园林绿化植物,还是水土保持、生态修复的良好植物材料。此外,榕属植物因与其传粉昆虫榕小蜂之间严格专一的互利共生关系,还是研究物种间协同进化关系的经典材料[8]。
种子是植物生活周期的一个重要阶段,种子萌发是植物生命的开始以及植物生活史的一个重要环节。植物种群周期中以种子形式出现的阶段称为潜在种群,种子萌发是植物从潜在的种群转变为现实种群的关键,这一阶段对种群个体的繁殖、扩展和抵御不良环境有着非常重要的意义[9]。不同的种子具有不同的萌发特性。学术界对多种经济植物、珍稀濒危植物的种子生物学特征开展了较多研究[10-12],但对榕属植物的种子萌发等特性开展的研究较少。目前大部分榕属植物还处于野外自然生长状态,对其了解和利用较为有限。另外由于热带地区橡胶树等经济作物的种植,使得榕树的野外种群遭到了人为砍伐等破坏,给热带雨林生态系统的稳定带来了负面影响。本研究以西双版纳地区4种榕属植物为研究对象,这4种榕树具有不同的生活类型,有幼时多附生的斜叶榕,有小乔木的对叶榕和木瓜榕,也有高大乔木的聚果榕。这4种榕树处于热带雨林群落演替的不同阶段且生境也不尽相同[13-14]。比较它们的种子萌发和幼苗生长发育特性,有助于进一步了解不同榕属植物的生物学特征,同时为榕树植物的园林栽培应用以及生态保护等提供科学依据。
1 材料和方法
1.1 试验材料
在位于云南省勐腊县勐仑镇的中国科学院西双版纳热带植物园及其周边地区,选定所需的4种榕属植物作试验用树,并对其进行跟踪观察。待其发育到成熟期时,采集其发育良好的成熟榕果,每种榕树采集10个。从榕果内挑出发育饱满、无病害、且大小较均匀的种子待用。4种榕属植物的属性见表1。
表1 4种榕属植物特征

种名亚属繁殖类型特征描述斜叶榕(Ficus tinctoria)糙叶榕亚属(Sycidium)雌雄异株果球形或球状梨形,单生或成对腋生,斜叶榕幼时多附生,小乔木,花果期为冬季至翌年6月。对叶榕(Ficus hispida)聚果榕亚属(Sycomorus)雌雄异株果腋生或生于落叶枝上,或老茎的下垂枝上,树为灌木或小乔木,多生长在沟谷潮湿地带,花果期为6—7月。木瓜榕(Ficus auriculata)聚果榕亚属(Sycomorus)雌雄异株果成簇生长于老茎短枝上或树干基部,乔木或小乔木,多生于低山沟谷潮湿雨林中,花果期为8月至翌年3月。聚果榕(Ficus racemosa)聚果榕亚属(Sycomorus)雌雄同株主干或老茎结果,高大乔木,喜生于潮湿地带,常见于热带雨林的溪边、河畔边,花果期为5—7月。
1.2 种子形态特征比较
种子形态比较:种子用纯水冲洗干净后用滤纸吸干,每种种子随机挑选30粒,利用LEICA(M 165 FC)正置显微镜对种子进行拍照,然后测量种子的长轴、短轴;种子质量观测:采用百粒法,从新鲜成熟榕果中随机抽取300粒健康、饱满的种子,在精密天平(OHAUS CP 114)上称量其鲜重和干重,换算种子千粒重;计算种子的表面积时,将种子视为椭圆形,则其表面积计算公式为:
表面积=π×种子长×种子宽/4。
1.3 发芽试验
将培养皿进行消毒,在每个培养皿内垫上约1 cm厚脱脂棉,然后在脱脂棉上铺上灭菌滤纸,用超纯水浸润脱脂棉和滤纸,以滤纸充分湿润但水分不浸出为准。试验用种子先放入超纯水中浸泡24 h,然后置于滤纸上。每皿均匀摆放1行种子共15粒,给予每粒种子编号以便后续记录,每个处理3次重复。将培养皿放置在赛福 PRX-350 C人工气候箱中。参考相关报道,将人工气候箱温度设置为30 ℃,光照为12∶12(L∶D),湿度为80%[15]。实验过程中每天检查培养箱,若滤纸干燥及时补充水分。试验持续6周,每天记录发芽的种子数(以胚根长度达到2 mm计算为发芽)和每粒种子的发芽时间,共16 d。每周使用宝工PD-151电子游标卡尺对培养皿中的每粒发芽种子进行茎叶长测量并记录。另外,从每种榕树种子中挑选饱满种子300粒,放入泰斯特101-1 AB干燥箱烘干至恒重。
表3 4种榕树种子的萌发指标比较

种名发芽率(%)发芽势(%)开始萌发时间(d)平均发芽速度发芽指数斜叶榕95.56±3.85a91.11±10.18a610.79±1.51a7.50±0.85a对叶榕91.11±10.18a77.78±7.70a611.08±0.73a10.00±2.39b木瓜榕75.56±10.18b28.89±21.43b612.95±0.68b4.33±2.41c聚果榕93.33±6.67a93.33±6.67a45.59±0.76c6.67±1.14d
1.4 指标测定
1.4.1 种子萌发特征测定
发芽率、发芽势、平均发芽速度、发芽指数公式为:
发芽率(%)=种子发芽数/供试种子总数×100%(实验最后1 d的发芽数);
发芽势(%)=n9/供试种子总数×100%;
式中:n9为种子日萌发最高峰第9天的发芽数;
平均发芽速度=∑(D×n)/∑n;
式中:D为置床时间起算天数;n为相应各天的发芽粒数;
发芽指数GI=∑(Gt/Dt);
式中:Gt为第t天的种子发芽数,Dt为相对应的种子发芽天数。
1.4.2 幼苗生长发育特征
连续培养6周,每周用电子游标卡尺(宝工PD-151)测定每株幼苗的胚根和胚芽长。培养结束后用电子天平称量每个种的幼苗鲜重。
1.5 数据分析
比较种子形态特征时,采用单因素方差分析(One-Way ANOVA),首先对数据的方差齐性进行检验:若方差齐,采用单因素方差分析中的多重比较(LSD);若方差不齐,先进行Log转换,后再次进行方差齐性检验,若方差齐采用单因素方差分析中的多重比较(LSD),若方差不齐则采用Dunnett’s T3检验(该检验未假定方差齐性)。采用卡方检验(Chi-square test)分析不同种子的发芽率和发芽势的差异。采用单因素方差分析比较不同种子的平均发芽速度、发芽指数以及茎长和根长。
计算数据采用平均值±标准误表达。采用Excel 2010软件进行数据整理和作图,采用SPSS软件进行数据分析。
2 结 果
2.1 4种榕树种子形态特征
4种榕树种子的长度存在显著差异(ANOVA,F(3,116)=124.96,p<0.001),其中斜叶榕、对叶榕和木瓜榕种子长度最大,且差异不显著,而聚果榕种子长度显著小于其他3种榕树种子(Dunnett T3,p<0.001)。种子宽度在4种榕树之间也存在显著差异(ANOVA,F(3,116)=97.00,p<0.001),且4个种的两两之间差异均显著(Dunnett T3,p<0.001),宽度大小为:对叶榕>木瓜榕>斜叶榕>聚果榕。4种榕树种子表面积同样存在显著差异(ANOVA,F(3,116)=179.56,p<0.001),其中对叶榕表面积显著大于其他3个种(Dunnett T3,p<0.001),斜叶榕和木瓜榕次之,二者无显著差异(Dunnett T3,p=0.975),聚果榕表面积最小(Dunnett T3,p<0.001)。种子千粒重表现为:斜叶榕>对叶榕=木瓜榕>聚果榕。4种榕树种子的含水量也表现出较大差异:木瓜榕>对叶榕>斜叶榕>聚果榕(表2)。
2.2 4种榕树种子萌发的生物学特征
4种榕树种子的发芽率表现出显著的种间差异(X2=11.25,p<0.05)。其中斜叶榕、对叶榕和聚果榕的萌发率都在90%以上,而木瓜榕萌发率(75.56%)显著低于其他3种榕树。发芽势和发芽率也呈现相同的结果,4种榕树的种子发芽势之间差异显著(X2=61.55,p<0.01)。其中斜叶榕、对叶榕和聚果榕之间差异不显著,而木瓜榕发芽势(28.89%)显著小于其他3种榕树。另外,4种榕树种子的平均发芽速度之间差异同样显著(ANOVA,F(3,117)=75.76,p<0.001):木瓜榕>对叶榕=斜叶榕>聚果榕。而发芽指数在4种榕树的两两间均有显著差异(ANOVA,F(3,117)=297.40,p<0.001):对叶榕>斜叶榕>聚果榕>木瓜榕。
表2 4种榕树种子形态及特征比较

种名种子长(mm)种子宽(mm)表面积(mm2)千粒重(g)含水量(%)斜叶榕1.11±0.10a0.67±0.08a0.59±0.10a0.468.93对叶榕1.11±0.08a0.81±0.08b0.71±0.09b0.3510.39木瓜榕1.06±0.04a0.72±0.04c0.60±0.04a0.3515.19聚果榕0.82±0.06b0.55±0.05d0.35±0.05c0.147.57
2.3 4种幼苗生长发育特性
从图1可以看出,4种榕树的茎生长呈现出一定差异,斜叶榕的茎生长高峰出现在第1周,且生长速度较快,第1周后就停止生长。而对叶榕、木瓜榕和聚果榕的生长高峰均出现在前2周,之后4周仍可缓慢生长一定长度,第6周时斜叶榕、对叶榕、木瓜榕和聚果榕的茎长分别为2.74、1.78、2.45 mm和1.93 mm,除对叶榕和聚果榕之间差异不显著外(LSD,p=0.45),总体呈现显著差异(ANOVA,F(3,176)=19.47,p<0.001)(图1)。

图1 4种榕树幼苗茎长发育比较

图2 4种榕树根长发育比较
4种榕树的根长也呈现出较大差异:其中对叶榕和聚果榕的根在生长2周后基本停止生长,而斜叶榕和木瓜榕的根长直到第6周仍然在持续生长。第6周时斜叶榕、对叶榕、木瓜榕和聚果榕的根长分别为33.4、14.88、35.03 mm和15.63 mm,总体间差异显著(ANOVA,F(3,176)=30.79,p<0.001),但斜叶榕和木瓜榕(LSD,p=0.52),对叶榕和聚果榕(LSD,p=0.84)之间差异不显著(图2)。此外,4种榕树幼苗的鲜重大小依次为:斜叶榕>木瓜榕>对叶榕>聚果榕(图3)。
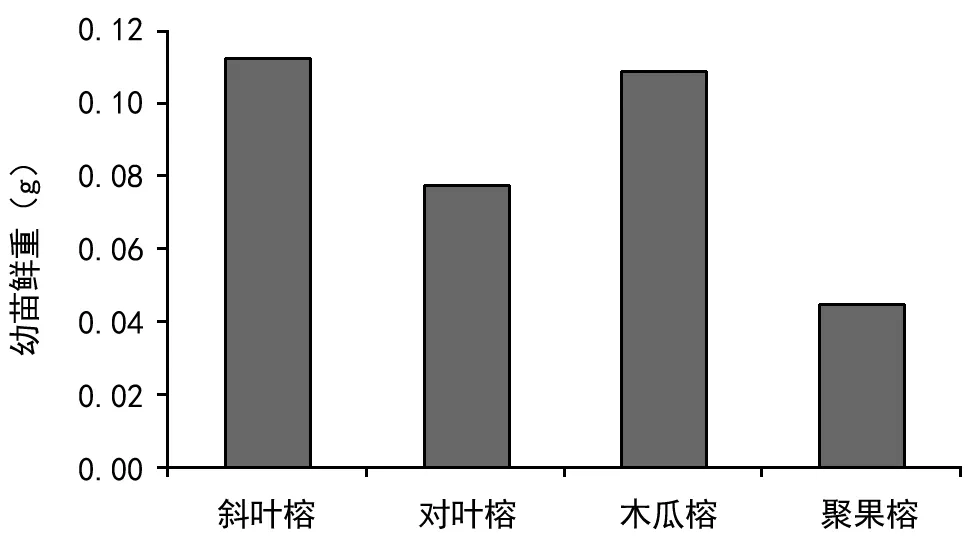
图3 4种榕树幼苗生物量比较
3 讨 论
榕属植物是国际上公认的热带雨林中的一类关键植物类群,榕树生活型有附生、半附生、攀援、蔓生灌丛等多种形态,在热带亚热带地区的森林、山地、溪边、城镇乃至石头缝中随处可见[7]。榕树也是我国热带亚热带地区城市常用的行道树、园景树和庭院树。由于榕属植物特殊的生理生态特点,使得其成为科学研究的理想材料。但目前对榕属植物种子萌发及幼苗建成的研究较少,对其进行研究,有助于了解不同种榕树在热带雨林生态系统中的角色,也有助于科学选择榕属植物作为园林树种。
研究表明:决定植物种子能否正常萌发以及决定植物萌发后能否继续发育成为健康植株,涉及到很多因素,但可概括为2个主因,即种子的内部生理条件和种子发芽的外部生态环境。种子萌发的内在生理条件往往容易被忽略,但实际上外因必须通过内因才起作用,其中种子的发芽力、整齐度,幼苗状况等,是种子萌发内在生理条件的直接体现。对濒危植物结实特性的研究表明,在天然条件下,种子向幼苗的转化普遍是濒危植物的脆弱环节[16]。本试验设计的萌发条件,是通过前期工作验证的榕树较理想的萌发状态,与西双版纳地区的自然环境相似。在统一的4种榕属植物外在生长条件的情况下发现:4种榕树植物的种子大小和重量均存在较大差异,对叶榕的种子最大,聚果榕的种子最小。一般来说,在抵御自然界严酷环境的能力上,体积较大的种子比体积小的种子能力更强,在幼苗建立阶段,大种子也比小种子更有优势[17]。所以从进化角度来看,生长在资源贫乏和严酷环境中的种子更容易进化出较大质量的种子。在本试验中,相较于其他3种榕树,聚果榕为热带喜水树种,一般生长在溪边或河边,生长环境的水热条件好,加之聚果榕一般为高大乔木,很少面临光照限制等情况[18],所以进化并不支持聚果榕生长出大种子来维持其幼苗的自养状态。
发芽率、发芽势和发芽指数从一定程度上代表了种子的生命力,木瓜榕的这3个发芽指标均低于其他种,说明木瓜榕的种子生命力较其他3个种差,在抵抗极端环境时的能力也相对较差,所以应谨慎选择木瓜榕这类树种作为行道树种。另外,对4种榕树幼苗生长阶段的测量数据表明,无论是茎还是根的生长,斜叶榕均最快。根茎的快速生长,有利于其根系快速深入土壤,得到更多的水分和养分,这也解释了为什么斜叶榕可以快速在环境中建立种群,成为先锋种[6,19]。以此相对应的是,聚果榕根茎的萌发速度均最慢,认为聚果榕的这一萌发特征可能与其种子较小有关。较缓慢的生长速度,不利于聚果榕在严酷环境中与其他树种竞争,这也从另一个侧面验证了聚果榕一般只生长在水热条件较好的生境中。
目前学术界对经济作物的种子研究较多,而对榕属植物的种子生长和幼苗建成等研究较少报导。实际上,榕树除了在热带雨林生态系统中扮演了关键角色外,还具有很高的开发利用价值,例如对叶榕和歪叶榕的皮可以提取单宁,聚果榕可以作为紫胶虫的优良寄主树,厚皮榕可以作为板材树种,木瓜榕、高榕等的嫩叶可作为食用蔬菜,叶榕、斜叶榕的根、树浆等可入药[7]。但目前对于榕树的利用方式较为粗放,多为直接获取野外资源,而对榕树的人工栽培和利用研究开展得较少。此外,在榕树作为行道树的选择上,也缺乏较系统的研究,导致在一些温带和亚热带城市,榕属行道树出现大规模冻害等情况[20]。榕属植物既是很好的绿化树种,也是具有潜力的经济植物,但对其利用必须建立在科学研究基础上,在今后的研究中,有必要在更大范围内对不同榕树的种子萌发、幼苗建成以及抗旱能力等开展系统研究,以利于榕属植物的科学利用。
猜你喜欢
趣味(语文)(2021年9期)2022-01-18
今日农业(2021年10期)2021-11-27
文苑(2020年11期)2021-01-04
学生天地(2020年16期)2020-08-25
学生天地·小学低年级版(2019年1期)2019-03-18
学生天地(2019年3期)2019-03-05
故事作文·高年级(2017年8期)2017-08-23
中国火炬(2015年12期)2015-07-31
红豆(2015年1期)2015-03-02
中国火炬(2013年3期)2013-07-24
