
香菇草匍匐茎及叶结构对不同水湿生境的生态适应性
2018-10-12 09:11:36缪丽华王莹莹乔东东杨琴琴季梦成
湿地科学与管理 2018年3期
缪丽华 王莹莹 乔东东 杨琴琴 季梦成*
(1 中国湿地博物馆,浙江 杭州 310013; 2 浙江农林大学,浙江 杭州 311300)
为了适应不同的生态环境,植物进化过程中器官的形态结构特征能明显表现出环境因子的影响,植物茎、叶在环境压力下形成各种适应类型,是对环境变化较敏感且可塑性较大的器官(Randy,2002)。形态结构改变越显著,植物对环境适应性越强(王桂芹等, 2011)。在干旱胁迫下,叶片的形态结构会发生变化,其形态结构的改变与植物的耐旱性有着密切的关系(Jiang et al, 2001),叶肉栅栏组织的细胞层数增加,密度增大,既可避免干旱地区强光照对叶肉细胞的灼伤,又能有效利用衍射光进行光合作用(薛静等,2010)。
表型可塑性指同一个基因型对不同环境变化而产生不同表型的能力,是植物适应自然选择的基本特征(解焱,2008)。表型的高度可塑性是植物与其生长环境长期相互作用的结果,也可体现外来植物的入侵特征(Rejmanek et al,2003)。许多外来入侵植物茎的形态结构对水分变化具有较强的可塑性(陶勇等,2004a; 赵相健等,2015),空心莲子草茎结构随水分条件的变化有显著差异,可在旱生性与水生性特点之间转换(陶勇等,2004b)。
香菇草Hydrocotyle vulgaris为伞形科天胡荽属多年生草本植物,原产欧洲及北美,作为水生观赏植物引入我国,在长江以南地区广泛应用。该种生态适应性较强,化感作用明显(杨琴琴等,2013),入侵风险较大(缪丽华等,2011;全晗等,2016)。目前,对香菇草的研究主要还涉及生理特性(闫道良等,2014)、生长繁殖(Liu et al,2017)、化感作用(杨琴琴等,2013)、水污染防治(金树权等,2010)、环境修复(Vafaei et al, 2013)等方面,未见香菇草解剖结构对水分变化的适应性、表型可塑性报道。
1 材料与方法
取材地点为浙江省杭州市西溪湿地国家公园内的湿地植物观测点,观测点坡度20°,沿池塘浅水水域向漫滩及陆地过渡,周边隔离不受外界干扰,丰枯水期水位差约10 cm。以池塘浅水水域作为水生生境,以池塘漫滩(丰水期浸润)作为湿生生境,全年未入水的陆地作为旱生生境。2011年4月定植,2012年10月取样,3种生境香菇草盖度均为100%。随机选取10株生长良好、无病虫害的香菇草植株,将叶片切割为3 mm×5 mm左右的片段,匍匐茎切割长度为5 mm左右,FAA固定液固定,按常规石蜡切片制作方法制成永久装片。制作香菇草鲜叶下表皮临时玻片标本,镜检计测气孔长度。
统计分析:采用EXCEL软件完成所有数据处理与分析。叶片结构紧密度CTR=栅栏组织厚度/叶片厚度×100%;叶片结构疏松度SR=海绵组织厚度/叶片厚度×100%;气孔指数(%)=单位视野气孔数/(单位视野普通表皮细胞数+单位视野气孔数)×100%。可塑性指数参考Valladares等(2000)的方法计算,即各个性状所测量的最大、最小值之差与最大值的比值,表示为PL=[max(i)- mix(i)]/max(i)×100%。
2 结果与分析
2.1 水分变化对香菇草叶表皮气孔的影响
随着生境水分含量的降低,香菇草叶片表皮气孔长度先增大后减小,气孔指数逐渐增大(表1)。湿生型气孔长度和气孔指数较水生型均无显著差异,旱生型气孔长度较湿生型降低了12.34%,气孔指数较湿生型增加了14.08%,差异均达到极显著水平(P<0.01,下同)。

表1 水分对香菇草叶表皮气孔的影响Table 1 Eあect of water condition on stoma of H. vulgaris
2.2 水分变化对香菇草叶片解剖结构的影响
对3种不同生境香菇草叶片的上、下表皮细胞厚度、栅栏组织厚度、两层栅栏细胞厚度比、海绵组织厚度、叶片厚度、叶片结构紧密度、疏松度、栅海比(栅栏组织与海绵组织厚度比值)9个参数进行比较分析(表2)。
单因素方差分析的结果表明,生境水分含量的变化对香菇草叶片结构的全部参数都有显著影响,其中对下表皮厚度的影响呈显著水平(P<0.05,下同),对其他参数的影响呈极显著水平。湿生型上、下表皮细胞厚度、栅栏组织厚度、叶片结构紧密度、栅海比较水生型均极显著增大,两层栅栏细胞厚度比和叶片厚度较水生型显著增大,湿生型海绵组织厚度、叶片结构疏松度较水生型均极显著减小,旱生型叶片上表皮细胞、海绵组织厚度、叶片疏松度、叶片厚度较湿生型均极显著减小,两层栅栏细胞厚度比、叶片结构紧密度、栅海比较湿生型均极显著减小,下表皮细胞厚度和栅栏组织厚度较湿生型差异不显著。
随着土壤含水量的降低,香菇草上、下表皮细胞厚度、栅栏组织厚度以及叶片厚度均表现出先增大后减小的趋势,湿生型较水生型增加比例依次为:栅栏组织厚度>上表皮细胞厚度>下表皮细胞厚度>叶片厚度;旱生型较湿生型减小比例依次为:上表皮细胞厚度>叶片厚度>下表皮细胞厚度>栅栏组织厚度。海绵组织厚度、叶片结构疏松度随生境含水量降低而减小,湿生型较水生型分别减小了7.64%和9.14%,旱生型较湿生型分别减小了53.77%和42.16%。两层栅栏细胞厚度比、叶片结构紧密度与栅海比随生境水分含量减少而增加,湿生型较水生型增加了6.94%、29.40%和42.33%,旱生型较湿生型分别增加16.98%、24.72%和115.54%。

表2 三种生境香菇草叶片解剖结构单因素方差分析结果Table 2 The result of One Way-ANOVA for eight morphological parameters of leaves on 3 populations of H. vulgaris
当生境水分减少到干旱状态时,叶片上表皮细胞缩小,形成紧密的表皮细胞层,能够有效防止水分散失。干旱型叶片结构紧密度与栅海比增大,较水生型和湿生型具有更紧密的栅栏细胞层,能够有效防止水分散失。叶片的解剖结构反映出香菇草具有一定的耐旱性。
2.3 不同生境水分含量对香菇草匍匐茎解剖结构特征的影响
香菇草匍匐茎结构主要包括表皮细胞层、厚角细胞层、薄壁组织、髓和维管束。生境水分含量变化对香菇草匍匐茎的厚角组织厚度和导管数有显著影响,对其他参数的影响均达到极显著水平。
旱生型匍匐茎表皮细胞厚度/匍匐茎直径的比值与湿生型没有显著差异,旱生型厚角组织厚度和薄壁组织厚度较湿生型显著减小,但厚角组织和薄壁组织在匍匐茎中所占比值较湿生型增加,有利于水分和养分的贮存。旱生型维管束数量较湿生型显著增加,两种生境匍匐茎的导管数量差异不显著(表3)。

表3 三种生境香菇草匍匐茎解剖结构单因素方差分析结果Table 3 The result of One Way-ANOVA for nine morphological parameters on 3 populations of H. vulgaris
香菇草匍匐茎表皮细胞厚度、厚角组织厚度、薄壁细胞厚度、髓直径、匍匐茎直径以及髓直径/匍匐茎直径的比值均随着生境水分含量的降低表现出先增大后减小的趋势,其中,湿生型较水生型增加比例最大的参数是髓直径,为82.99%,其次分别为薄壁组织厚度、匍匐茎直径、髓直径/匍匐茎直径、表皮细胞厚度和厚角组织厚度;各参数旱生型较湿生型减小比例依次为:髓直径>表皮细胞厚度>匍匐茎直径>薄壁组织厚度>髓直径/匍匐茎直径>厚角组织厚度。维管束数和导管数随生境水分含量的降低逐渐增加,湿生型较水生型增加比例分别为25.53%和81.25%,旱生型较湿生型增加比例分别为6.78%和17.24%。茎中气孔长度随生境水分含量的降低逐渐减小,湿生型较水生型减小了34.50%,旱生型较湿生型减小了43.96%。
2.4 不同生境下香菇草各解剖结构可塑性指数分析
2.4.1 叶片解剖结构可塑性指数分析 不同水分生境下香菇草叶片解剖结构特征值的可塑性指数见表4。
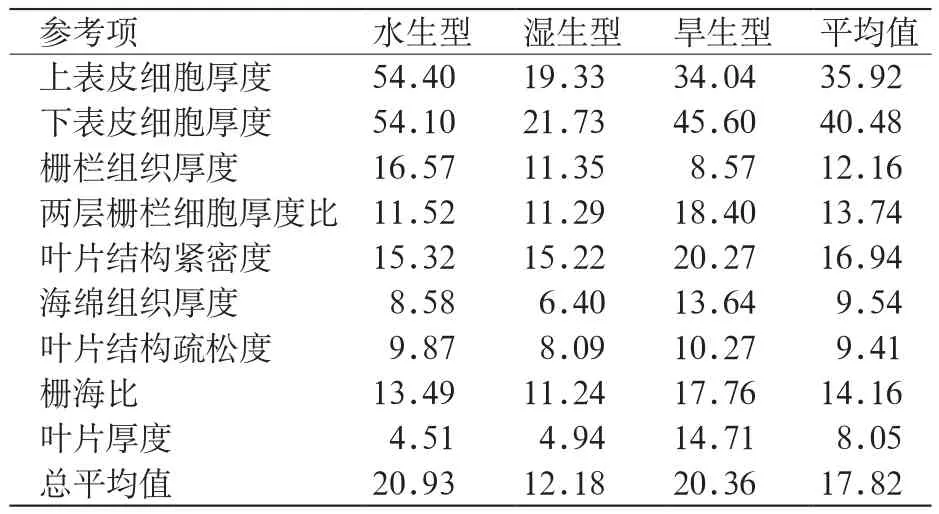
表4 三种生境香菇草叶片解剖结构可塑性指数 %Table 4 Elasticity index for eight morphological parameters of leaves on 3 populations of H. vulgaris %
3种生境中香菇草叶片可塑性指数总平均值大小排序为:水生型>旱生型>湿生型。其中,水生型上、下表皮细胞厚度和栅栏组织厚度可塑性指数较湿生型和旱生型高。两层栅栏细胞厚度比、叶片结构紧密度、海绵组织厚度、叶片结构疏松度、栅海比和叶片厚度可塑性指数为旱生型最高。香菇草下表皮细胞厚度可塑性平均值较上表皮高,叶片结构紧密度的可塑性平均值较疏松度参数高。在各参数可塑性指数中,叶片厚度的平均值最小,其次是叶片结构疏松度和海绵组织厚度,其可塑性指数平均值均不超过10%。
2.4.2 匍匐茎解剖结构可塑性指数分析 不同水分生境下香菇草匍匐茎各解剖结构可塑性指数见表5。匍匐茎各参数中导管数的可塑性指数最高,其次为气孔长度,可塑性指数最低的为匍匐茎直径。3种生境香菇草匍匐茎可塑性指数强弱依次为:水生型>湿生型>旱生型,但平均可塑性指数差异不明显。就不同生境而言,水生型香菇草匍匐茎气孔长度、匍匐茎直径的可塑性指数分别为最高(56.55%)和最低(5.05%);导管数可塑性指数最高均出现在旱生型(85.71%)和湿生型(60%),湿生型和旱生型匍匐茎直径的可塑性指数最低,分别为2.85%和3.37%。
3 结论与讨论
植物长期在某一特定环境生存,必然形成适应该环境的形态结构以保证植株的正常生长。叶片作为植物重要的营养器官,受到环境中光照、温度、水分等因素的影响较大,是植物进化过程中对环境变化较敏感且可塑性较大的器官(李芳兰等,2005)。于海秋等(2003)对水分胁迫下玉米气孔变化的研究发现,经水分胁迫的玉米叶片气孔密度增大, 气孔的长、宽明显减小。香菇草叶片表皮气孔长度减小,气孔指数增加,旱生型叶片小而多的气孔更有利于减少水分散失,与前人的研究结果一致。
叶片上表皮受到阳光直接照射,在长期干旱的环境下,叶片会形成小而密的表皮细胞以减少水分的散失,从而提高水分利用率(Bosabaalidiset al,2002),旱生植物的叶肉向着提高光合作用效率的方向发展,栅栏组织高度发达,海绵组织退化甚至缺失(燕玲等, 2000)。本试验中,随着生境水分含量的降低,香菇草叶片厚度减小,栅栏组织与海绵组织比增大,栅栏组织细胞层显著增厚。在香菇草叶片湿生型与水生型各参数中,栅海比受生境水分含量变化影响最大,其次是栅栏组织厚度;在香菇草旱生型与湿生型各参数中,受水分含量变化影响最大的分别是栅海比和海绵组织厚度,叶肉组织厚度变化是香菇草叶片应对生境水分含量变化最主要的方式。随着生境水分含量的降低,匍匐茎的直径先增大后减小,气孔长度减小,导管数量增加,有利于干旱环境下香菇草植株地上、地下部分之间以及各植株之间水分的运输。叶片和匍匐茎的解剖结构变化显示香姑草对水湿生及干旱环境具有良好的适应性,其结构变化趋势取决于环境的水分条件。

表5 三种生境香菇草匍匐茎解剖结构可塑性指数 %Table 5 Elasticity index for eight morphological parameters of runners on 3 populations of H. vulgaris %
表型可塑性是衡量香菇草对不同水分条件适应能力的重要指标,表型可塑性高低常与其对环境的潜在适应能力大小紧密联系。香菇草叶片上、下表皮细胞厚度都具有较高的可塑性,而栅栏组织厚度、海绵组织厚度和叶片厚度可塑性指数较低,说明直接接触环境的叶片表皮细胞层对环境的变化反应最为迅速。湿生型叶片结构各项可塑性指数均较低,结构更加稳定,可见较水生型和旱生型生境,香菇草更适宜于湿生型生境。

香菇草(作者供图)
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
落叶果树(2021年6期)2021-02-12 01:29:10
体育科技(2016年2期)2016-02-28 17:06:07
江苏农业科学(2015年8期)2015-09-10 15:54:14
西南医科大学学报(2015年1期)2015-08-22 13:01:48
生态学报(2015年4期)2015-03-10 12:19:17
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
生态学报(2015年23期)2015-02-11 02:01:41
