
小麦抗逆相关基因 TaGB1的克隆及其表达特性分析
2018-10-09 02:33王小婷王文静杨雯晶胡卫国
麦类作物学报 2018年9期
王小婷,王文静,李 波,杨雯晶,胡卫国
(1.河南省农业科学院小麦研究所/河南省小麦生物学重点实验室/农业部黄淮中部小麦生物学与遗传育种重点实验室/国家小麦工程实验室,河南郑州 450002;2.西北农林科技大学农学院/旱区作物逆境生物学国家重点实验室,陕西杨凌 712100;3.陕西省农业机械鉴定推广总站,陕西西安 710003)
异源三聚体GTP结合蛋白(G蛋白)是由α、β、γ亚基(Gα、Gβ、Gγ)组成的膜蛋白复合物,参与介导真核生物中保守的信号传导网络[1]。在传统模式中,异源三聚体GTP结合蛋白通过细胞膜限定结合外界的多种信号,然后通过G蛋白偶联的受体蛋白(GPCR)将外界信号传递至下游的效应蛋白,最终引起转录水平的调控,进而响应外界环境的刺激。
通常认为植物G蛋白信号传导机制与动物G蛋白相似,但是植物中GPCR信号元件的种类和数量与动物中不同[2]。在哺乳动物中,由20个Gα、5个Gβ和12个Gγ亚基组成的异源组合为超过800个基因编码的巨大的GCPRs家族提供了信号特异性[3]。哺乳动物G蛋白偶联的信号通路参与体内大量的生理过程,并且跟许多疾病状态相关,这使得G蛋白成为药物治疗的重要靶标[4]。除参与疾病治疗之外,G蛋白在发育阶段的信号传递过程中也履行重要的功能[3],如有丝分裂过程中的保守功能[5]。然而与哺乳动物及其他真核生物相反,植物中仅有几种常见的G蛋白亚基种类,如模式植物拟南芥基因组仅含有1个Gα、1个Gβ和2个Gγ亚基[6]。除此之外,拟南芥基因组不具备编码动物中已知效应蛋白同系物的能力[7]。在水稻基因组中,含有1个Gα( RGA1)[8]、1个Gβ(RGB1)[9]和目前已报道的Gγ1(RGG1)[10]。与哺乳动物相比,植物的G蛋白相关组件较少[7],但诸多研究表明,植物G蛋白同样参与众多的生理过程,包括细胞分裂[11-12]、离子通道调节[13-14]、种子萌发[15]、生物和非生物胁迫响应[16-17]、蓝光介导的响应[18]等。
小麦是世界两大谷类作物之一,同时也是能最大限度适应恶劣环境条件的主要粮食作物[19]。目前,关于异源三聚体G蛋白在普通小麦中的研究报道多是关于抗锈病、白粉病或蚜虫等[20],且研究多是关于异三聚体G蛋白中的小G蛋白[21-24]。从普通小麦品种S615中,研究人员克隆得到编码G蛋白α亚基的两个基因TaGA1和TaGA2[25]。编码G蛋白β亚基基因的全长cDNA序列也从S615中被克隆得到,被命名为 TaGB1,其编码380个氨基酸,并且与其他植物和动物的G蛋白β亚基结构相似, TaGB1同样也具有7个WD40保守域[26]。但以上研究并未就 TaGB1的基因特性及表达情况进行分析,其与双子叶植物同源基因的进化关系等方面研究也存在空白。
本研究选取典型的双子叶模式植物拟南芥为比较对象,分析小麦G蛋白β亚基基因 TaGB1与其他植物中的G蛋白β亚基之间的亲疏关系,通过系统发育进化树研究 TaGB1进化上的亲缘关系,并分析 TaGB1在不同胁迫处理条件下的表达情况、蛋白定位以及组织特异性等,以期为解析小麦中G蛋白β亚基的功能奠定基础。
1 材料与方法
1.1 试验材料
小麦品种济麦22种子由本实验室保存,大肠杆菌感受态细胞TOP10购于江苏康为生物科技有限公司,亚细胞定位表达载体16318h-GFP由本实验室保存。
1.2 方 法
1.2.1 小麦材料处理
取适量小麦种子于培养皿内,7‰过氧化氢浸泡12 h破除休眠,清水冲洗3遍,室温萌发后移至96孔透明板进行水培,在温度22 ℃、相对湿度65%、光照周期16 h/8 h下生长10 d左右。整株及根、茎、叶分别取样后,液氮速冻,-80 ℃保存。
取水培10 d左右的生长状态一致的小麦幼苗进行干旱、ABA、盐及热胁迫处理。干旱胁迫处理:用吸水纸擦干幼苗表面水分,置于干燥滤纸上对幼苗进行自然脱水处理,分别于0 h、2 h、3 h、12 h、24 h取样。 ABA胁迫处理:小麦幼苗置于含有100 μmol·L-1ABA的水培液中进行胁迫处理,分别于0 h、2.5 h、7 h、25 h、48 h、72 h不同时间点整株取样。100 mmol·L-1NaCl胁迫处理:取幼菌于含有100 mmol·L-1NaCl的水培液中培养,分别在0 h、1 h、4 h、12 h、24 h、48 h整株取样;热胁迫处理:除幼苗置于37 ℃温度下培养,分别在0 h、3 h、6 h、12 h、24 h、36 h整株取样,液氮速冻后-80 ℃保存。
1.2.2 总RNA提取及cDNA第一链的合成
植物样品总RNA提取按照全式金TransZol Up说明书操作步骤,1.5%琼脂糖凝胶电泳检测RNA质量,反转录cDNA第一链的合成参照全式金反转录试剂盒操作(货号:AT-311)。
1.2.3 小麦 TaGB1基因克隆
根据小麦G蛋白β亚基 TaGB1(ACCESSION:AB090160)序列,设计正向引物(5′-ATGGCGTCCGTGGCGGAGCTCAA-3′)和反向引物(5′-TCAGACTATCTTGCGGTGTCCAC-3′),以反转录得到的cDNA为模板,利用高保真DNA聚合酶PrimeSTAR进行PCR扩增。反应体系:2×PrimeSTAR GC Buffer 25 μL,dNTP Mixture(2.5 mmol·L-1)4 μL, 正向引物(10 μmol·L-1)1 μL,反向引物(10 μmol·L-1)1 μL,cDNATemplate 2 μL,PrimeSTAR HS DNA Polymerase(2.5 U·μL-1)0.4 μL,ddH2O 16.6 μL。PCR程序设定为95 ℃ 10 min;95 ℃ 45 s,69 ℃ 30 s,72 ℃ 1 min 30 s,35 cycles;72 ℃ 10 min;16 ℃保存。1.5%琼脂糖凝胶检测PCR产物大小,将目的片段利用TaKaRa MiniBESTAgarose Gel DNA Extraction Kit进行胶回收,构建至零背景克隆载体pZero Back/Blunt Vector,连接产物转化至感受态TOP10,挑取阳性单克隆菌液送北京奥科鼎盛生物技术有限公司测序。
1.2.4 生物信息学分析
利用分子生物学软件Clustal X 2.0和DNAMAN进行多重序列比对;运用在线网站ProtParam(http://web.expasy.org/cgi-bin/protparam/protparam)预测蛋白分子量和等电点;利用MEGA 6.0软件构建系统发育进化树;采用在线工具SMART(http://smart.embl-heidelberg.de/)进行蛋白质保守域预测。
1.2.5 TaGB1亚细胞定位
对亚细胞定位载体16318h-GFP进行BamHI单酶切,酶切产物回收,按照In-Fusion快速克隆试剂盒说明书将 TaGB1目的片段与酶切产物连接,连接产物转化至感受态TOP10,选取阳性菌液送测序。
选取生长1周的济麦22小麦幼苗叶片制备原生质体,采用PEG介导的转化方法将已测序正确的重组质粒16318h-GFP:: TaGB1和空载体同时转化至小麦原生质体细胞,避光培养16 h后观察蛋白的表达。
1.2.6 TaGB1基因的表达模式分析
分别以小麦幼叶、根、茎,以及不同胁迫处理条件下的小麦植株样品为材料,进行总RNA提取及cDNA第一链的合成,具体方法参照1.2.2中所述步骤。实时荧光定量PCR具体操作步骤依据天根Real Master Mix(SYBR Green)说明书,以反转录的cDNA为模板,内参基因FP(5′-CACTGGAATGGTCAAGGCTG-3′)和RP(5′-CTCCATGTCATCCCAGTTG-3′),依据 TaGB1序列设计引物分别为F1(5′-ACCCGCTTGATT ACAGGCTC-3′)、R1(5′-TGATATGTCCGAA CTGCCCG-3′)和F2(5′-CCCTGATGGTCAG AGGTTCG-3′)R2(5′-AGCGTGTCCCACACAT AACA-3′)。反应体系:2×SuperReal PreMix Plus 10 μL,50×ROX Reference Dye 0.4 μL,正向引物(10 μmol·L-1)0.3 μL,反向引物(10 μmol·L-1)0.3 μL,cDNA模板 2 μL,RNase-free Water 7 μL。
采用荧光定量PCR仪(ABI 7500)3步法反应程序进行PCR扩增:95 ℃ 15 min;95 ℃ 10 s,60 ℃ 20 s,72 ℃ 30 s,40 cycles,在72 ℃ 30 s时进行荧光信号采集。根据各样品特定荧光阈值下的Ct值,依据计算公式2-△△Ct比较基因的相对表达量。每个样本设置至少3个生物学重复。
2 结果与分析
2.1 小麦 TaGB1的基因序列特性分析
对济麦22的 TaGB1进行克隆(图1)、测序和BLAST分析表明,该基因分别位于小麦基因组4A、4B和4D染色体上,分别命名为 TaGB1A、TaGB1B和 TaGB1D。 TaGB1基因编码区全长1 143 bp,包含6个外显子和5个内含子,编码380个氨基酸,预测分子量为41 kD。 TaGB1在A、B和D染色体组间十分保守,对 TaGB1A、 TaGB1B和 TaGB1D多肽序列进行比对发现,其氨基酸序列同源性高达99.91%,仅在第330位氨基酸存在差异(图2),A染色体组中对应的氨基酸是缬氨酸(Val),而B和D染色体组中均是甲硫氨酸(Met)。由此推测 TaGB1在A、B和D染色体组中具有相似的功能。
M:DL2000;1~3: TaGB1 PCR 产物。M:DL2000; 1-3: TaGB1 PCR product.

红色标注为甲硫氨酸,绿色标注为缬氨酸。The red mark represents methionine,the green mark represents valine.
对 TaGB1的氨基酸序列进行结构域预测,发现其含有7个WD40保守域(图3)。为进一步分析 TaGB1与双子叶模式植物拟南芥的G蛋白β亚基 AGB1氨基酸序列差异,将 TaGB1各拷贝的多肽序列与 AGB1的多肽序列进行多重比对,结果(图4)显示, TaGB1与 AGB1同源性高达93.46%,且 TaGB1与 AGB1的第Ⅰ、Ⅲ、Ⅳ、Ⅵ和Ⅶ个WD40保守域相同,第Ⅱ个和第Ⅴ个WD40保守域存在差异。据此推断,小麦基因 TaGB1与 AGB1在功能上可能有部分相似性。
2.2 TaGB1系统发育进化分析
系统发育进化分析(图5)表明,不同植物的G蛋白β亚基蛋白序列高度相似, TaGB1在亲缘关系上与节节麦(Aegilopstauschii)、大麦(HordeumvulgareL.)、二穗短柄草(Brachypodiumdistachyon)、燕麦(AvenafatuaL.)等禾本科植物较近;从大的进化关系来看,G蛋白β亚基在物种进化上趋向于划分为单、双子叶两条途径,小麦G蛋白β亚基 TaGB1进化上属于单子叶植物大类,与拟南芥等双子叶植物的亲缘关系较远。

图3 TaGB1蛋白序列保守域(conserved domains)分析

图中红色横线标注为Ⅰ-Ⅶ7个WD40保守域,红色矩形标注为 TaGB1不同拷贝间氨基酸序列差异。Seven WD40 conserved domains in AGB1 are underlined in red,and sequence differences are indicated with red rectangle.
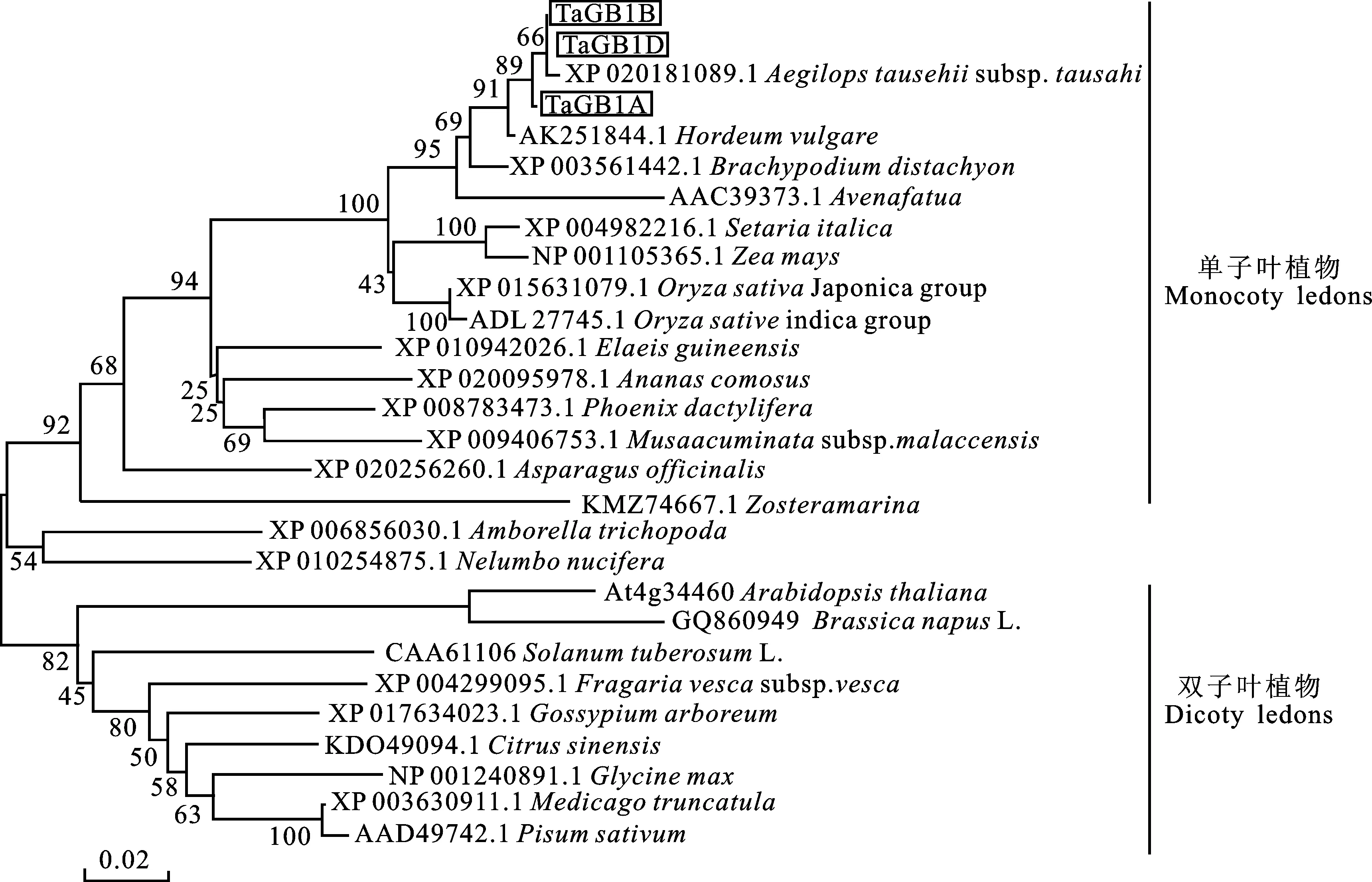
图5 系统发育进化树分析
2.3 TaGB1表达模式分析
检测结果(图6)表明,在ABA和NaCl胁迫下, TaGB1的表达趋势相似,在一定时间范围内,其相对表达量趋于上升,分别在处理后7 h和24 h左右达到最大值,之后趋于下降;在干旱胁迫下,在处理后0~12 h之间, TaGB1的表达基本上维持较稳定的水平,处理24 h左右后表达量迅速上升,约是处理前的3倍;热胁迫下, TaGB1的相对表达量在处理6 h左右时达到最大值,约是处理前的20倍。这说明 TaGB1基因表达受外界胁迫如ABA、NaCl、热和干旱诱导,推测该基因参与植物抗逆信号传导途径。
2.4 TaGB1亚细胞定位及组织特异性分析
以正常生长1周的小麦幼苗叶片为材料制备小麦原生质体,将重组质粒16318h-GFP:: TaGB1和空载体16318h-GFP同时转化原生质体细胞。观察结果(图7A)表明,阳性对照16318h-GFP在细胞膜、细胞质和核均能观察到荧光信号,证明PEG介导转化过程正常, TaGB1蛋白定位于胞质内成斑点状,且斑点状信号很强,除此之外,质膜上也存在微弱的荧光信号。组织特异性表达水平(图7B)显示,在正常生长条件下, TaGB1在幼苗的叶、根及茎中都有转录活性,但不同部位中相对表达水平存在差别,叶片中较高,而在根和茎中较低,说明该基因在感知外界信号和信号传递方面发挥作用。

图6 TaGB1表达模式
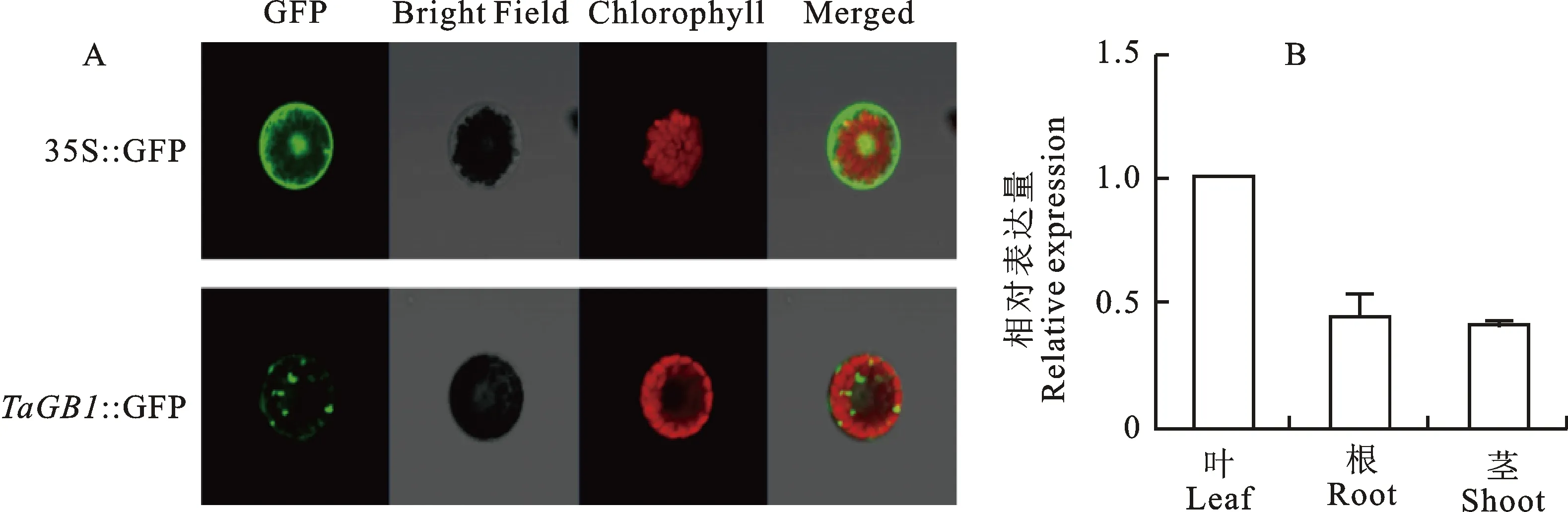
图7 TaGB1蛋白亚细胞定位及组织特异性分析
3 讨 论
研究表明,G蛋白β亚基C端含有一组7个重复串联的WD40结构[9],7个重复串联的WD40结构含有与Gα亚基以及下游效应因子相互作用位点,影响蛋白与蛋白之间的相互作用[35]。前人研究发现,编码小麦G蛋白β亚基的 TaGB1包含7个WD40保守域,与其他植物和动物的G蛋白β亚基类似[26],本研究结果与此一致。拟南芥有很多蛋白都包含WD40重复结构(WD40保守域),G蛋白β亚基 AGB1就是其中之一。本研究将 TaGB1的氨基酸序列与 AGB1的氨基酸序列进行多重比对,结果表明,除第Ⅱ个和第Ⅴ个WD40结构存在差异外,其余5个WD40保守域完全相同,说明G蛋白β亚基在不同物种的进化中具有高度保守性,WD40蛋白在多种细胞发育过程中发挥关键作用,因此,推测小麦抗逆基因 TaGB1可能与 AGB1具有部分相似功能。
TaGB1在亲缘关系上与禾本科等单子叶植物较近,而与拟南芥、烟草等双子叶植物较远,即分化形成两大分支(图5),这在植物G蛋白β亚基的相关研究中鲜有报道。根据上述 TaGB1和 AGB1的WD40保守域差异结果,推测这种进化上的分化与WD40保守域的差异存在一定的联系,但具体的机制仍需进一步探究。
AGB1的表达几乎贯穿植物生长的全部阶段,并且存在于各种组织器官中,包括根、花、茎、莲座叶、种子等[27-28]。本试验对 TaGB1进行表达分析表明, TaGB1在小麦根、茎、叶均有不同程度的表达,这与已有报道一致,但 TaGB1在叶中的表达量较高,原因可能是植物主要利用叶片感知外界刺激并传导至细胞内部引起一系列级联反应,或者可能是在植物不同的生长发育阶段中,不同组织部位依据各自发挥功能的主、次而引起表达量的变化,如水稻中 RGA1和 RGB1的mRNA在根部的积累低于在d线区域的积累,但是拟南芥中正好相反,即 GPA1和 AGB1的mRNA在根部的积累较高[29]。
研究表明,高温、氧化胁迫、高盐处理下,豌豆G蛋白成员PsGα和PsGβ在转录水平大量表达[31];水稻G蛋白各亚基受非生物胁迫和激素ABA诱导表达[32]。Colaneri等[33]和Ma[30]研究发现,G蛋白调节盐处理下植物的生长发育,并且拟南芥G蛋白β亚基突变体agb1-2对缺水的适应性增强[34]。 AGB1通过调节渗透胁迫、离子稳态和氧胁迫来正向调控植物的耐盐性[30]。本研究中, TaGB1在盐胁迫下上调表达,胁迫处理约24 h后表达量最高,随着时间的延长,表达呈现下降趋势, TaGB1在热胁迫下表达上调,处理后6 h左右表达量最高,这与已有的一些研究结果相符。环境胁迫会刺激植物产生内源ABA,这些ABA的产生调节植物的生长发育[35]。本研究中 TaGB1受外源激素ABA及干旱胁迫条件诱导上调表达,在处理后24 h达到最大值,说明 TaGB1 可能通过依赖ABA的信号通路调控植物的耐旱性。
有文献报道,其他植物中G蛋白β亚基的定位多是在质膜和核[41],以及细胞内其他的组分上[37-39]。本研究中,小麦 TaGB1蛋白主要定位于胞质,呈斑点状,质膜上亦存在微弱的GFP信号。关于小麦G蛋白β亚基胞质内斑点状定位,推测可能是高尔基体,准确定位有待进一步研究。
WD40重复保守域在植物体内扮演重要角色,包括介导不同蛋白质间的相互作用[2,40],以及参与蛋白质的降解过程等[41-42]。本研究中对 TaGB1的系统发育进化树分析以及 TaGB1和AGB1的WD40保守域差异分析,推测不同物种间G蛋白β亚基表达部位的差异可能与其进化分类相关,小麦G蛋白β亚基胞质内斑点状定位也可能与WD40功能域存在某种关联,因此关于 TaGB1斑点状定位具体的原因以及与WD40功能域之间的关系,仍需更进一步的研究。
猜你喜欢
今日农业(2022年16期)2022-11-09
麦类作物学报(2022年3期)2022-05-19
临床与实验病理学杂志(2022年3期)2022-04-06
金桥(2021年10期)2021-11-05
今日农业(2021年13期)2021-08-14
园艺与种苗(2020年12期)2021-01-08
世界最新医学信息文摘(2020年68期)2020-12-25
作文小学中年级(2020年4期)2020-06-11
烟台大学学报(自然科学与工程版)(2020年1期)2020-02-08
飞碟探索(2015年9期)2015-11-05
