
辽东栎幼苗根系形态特征对环境梯度的响应
2018-09-28 07:28关晋宏张文辉
生态学报 2018年16期
邓 磊,关晋宏,张文辉,3,*
1 西北农林科技大学林学院, 杨凌 712100 2 青海大学农林科学院, 西宁 810016 3 西北农林科技大学陕西省林业综合实验室, 杨凌 712100 4 青海高原林木遗传育种重点实验室, 西宁 810016
林木幼苗生长不仅取决于林木本身的遗传特性,而且在很大程度上受所处环境的影响[1- 2],这就导致幼苗形态具有很强的空间异质性,这种差异反映了幼苗对环境的适应能力[3- 5]。幼苗形态变化是植物适应环境的一种基本形式,严酷条件下幼苗首先通过形态变化来调整生存策略[6- 7]。不同纬度的环境因子表现出梯度性变化,这为研究林木器官形态对环境的响应提供了良好条件[8- 9]。根系形态特征是植物根系研究的基础内容,根系形态与年龄、生境和外界干扰等因素有关[10- 13]。由于植物地下部分具有较大的表面积、较高的生理吸收功能、较强的呼吸速率和较短的寿命,在植物生长发育和森林生态系统养分循环过程中发挥着重要作用[14]。根系在发育过程中形成明显的形态结构,可以用总长度、表面积、总体积、根尖数、分岔数、平均直径、比根长和组织密度等指标来进行描述,这些特征指标具有高度的形态异质性[15]。
辽东栎(Quercusliaotungensis)为壳斗科(Fagaceae)栎属(Quercus)植物,是秦岭北坡和黄土高原重要的地带性森林植被群落[16- 17]。研究辽东栎幼苗根系形态沿环境梯度的差异可以更好地反映辽东栎种群对环境的适应特性。例如,从秦岭北坡到黄土高原中部,生态因子、林分因子和土壤特征的差异,对辽东栎幼苗根系发育产生复杂的影响。有研究表明,植物可通过生物量分配来适应环境胁迫,干旱环境下植物采用减小主根直径的生物量分配模式,同时增加侧根数量来应对环境的胁迫[18];水分胁迫也会影响辽东栎根系生物量的分配[19]。国内外学者对辽东栎群落的结构[20- 21],生态功能[22],群落更新[23]以及土壤生态过程[24]等进行了广泛的研究,这些研究往往在单一的地理区域上开展,却很少有不同分布区气候、资源差异对幼苗根系的影响。特别是不同地理区域,幼苗根系形态会对环境做出怎么样的响应?这些过程是普遍发生的,还是与幼苗年龄有关?不同年龄的幼苗在根系形态、生物量方面存在差异[25],因此,本研究选择辽东栎分布的不同地理区域,开展辽东栎幼苗根系形态特征的研究,探讨辽东栎幼苗根系形态与环境因素的关系,为阐明辽东栎幼苗根系形态对不同地区的生态适应性提供依据,服务于区域植被管理。
1 研究地区与方法
1.1 研究区概况
陕西省地跨温带、暖温带和亚热带3个气候带,是我国辽东栎资源(分布面积和蓄积)较大的省份之一。本研究在陕西省境内以辽东栎为主要建群种的秦岭北坡太白山自然保护区、黄土高原南部黄龙山和黄土高原中部延安市宝塔区公路山作为研究地点,3个地区属于不同气候区域,依次为秦岭山地湿润气候区、渭北-延安高原丘陵沟壑半湿润气候区和延安-长城高原丘陵沟壑半干旱气候区。从秦岭北坡到黄土高原中部,形成了由温暖湿润到半干旱的逐步过渡,气候由南向北呈规律性变化(图1)。从秦岭北坡到黄土高原中部不同地区的辽东栎林样地概况见表1。
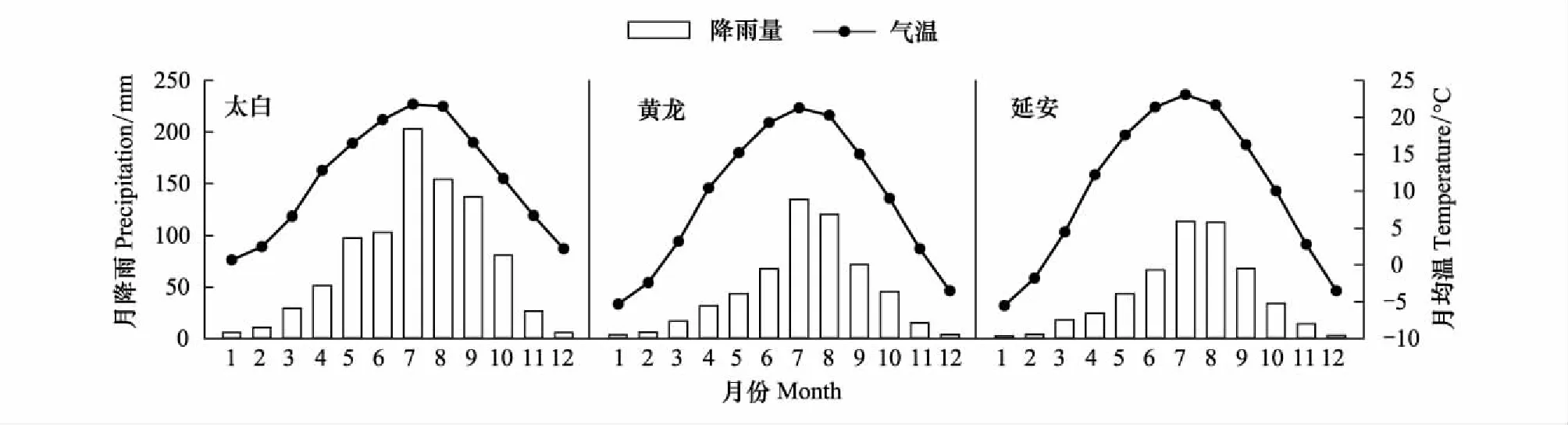
图1 研究地区多年平均各月降雨量和气温表变化(1971—2010年)
经过单因素方差分析,同行数据字母相同表示无显著差异,字母不同表示有显著差异(P<0.05)
1.2 研究方法
1.2.1 样地设置
样地的调查工作在2016年8—9月进行。分别在秦岭、黄龙、延安3个地区,选择生长较好林龄相近的辽东栎纯林布设样地。本研究充分考虑不同群落组成、生境条件和人为破坏等因子的代表性,在样地选择上采用一致的方法。针对种群分布特点,经过充分的实地踏查,在3个地区阴坡设置样地。样地布设过程中,遵循代表该区域辽东栎资源状况的基本原则,并兼顾样地与生境划分相结合的方法进行样点布设,样地间直线距离在300 m以上。每个地区设置样地3块,共完成样地9块。样地内分别沿对角线设置5个灌木样方和5个草本样方。样地面积20 m×20 m,灌木样方5 m×5 m,草本样方1 m×1 m。其中,样地地形、地貌、海拔和地理坐标用MagellanGPS315测定;坡向和坡度采用手持罗盘测定;土壤类型根据Food and Agriculture Organization (FAO)标准目测确定;土壤大于1 mm石砾含量依据土壤塑性的强弱确定,并采集样品带回实验室用“吸管法”验证;林龄利用生长锥(直径4.5 mm, 长500 mm)对样地内胸径大于平均胸径的10株辽东栎乔木进行钻芯取样,确定乔木年龄。
1.2.2 幼苗年龄的确定
将辽东栎整株幼苗全部挖出,做好标记。尤其在较大幼苗的挖掘中,利用小铲、毛刷等工具顺着侧根延伸方向轻拨根系,直至挖掘到幼苗根系末级,尽量避免临近其他植株根系干扰和幼苗损伤带来的测量误差。幼苗年龄通过枝条颜色、皮孔、芽鳞痕来确定。从枝条顶开始,向幼苗基部延伸,当年生枝条为一年生枝条;再向下根据芽鳞痕、枝条颜色依次确定出二年生、三年生、四年生和五年生枝条。按照本方法确定幼苗年龄,再用枝条基部年轮数验证[26- 27]。
1.2.3 幼苗根系指标的测定
将采集到的1—5年生辽东栎幼苗从幼苗基部切下,分为地上、地下两部分。地下部分根系测定时,先将根系取出后轻轻抖去粘在根系上土壤,立即用蒸馏水冲去根表面黏附的土壤,平铺在干净滤纸上,以吸去多余水分。用Epson数字化扫描仪扫描不同地区的不同年龄根系,并将扫描后的图像存入计算机,用与扫描仪配套的WinRHIZO根系图像分析系统软件对各处理下根形态指标进行定量分析。扫描后将各龄级幼苗地上部分和地下部分分别装入信封内,放入85℃烘箱烘至恒重,即可得到各龄级幼苗地上、地下部分生物量。主要分析根系总长度、表面积、总体积、根尖数、分叉数、平均直径、比根长和组织密度等指标。每个地区不少于重复测量8株年龄一致的幼苗个体,共采集幼苗189株。测定数据按同一地区进行平均。
根据根系图像分析系统软件计算出的根系形态指标以及所测定细根生物量,可计算出不同地区辽东栎幼苗细根的比根长和组织密度。计算公式如下:
比根长(m/g)=根系总长度(m)/根系生物量(g)
组织密度(g/cm3)=根系生物量(g)/根系总体积(cm3)
1.2.4 土壤水分养分的测定
土壤表层养分的取样是沿样地内对角线各取3个0—20 cm土壤混合样品。其中土壤水分含量测定采用烘箱(85℃)法;土壤pH值用ZD-2型电位滴定计测定;有机质用重铬酸钾-H2SO4容量法进行测定;全N(半微量氏法),全P(钼锑抗比色法),速效P用NaHCO3浸提-钼蓝比色法;速效N用碱解扩散法[28- 29];速效K采用火焰光度法[30- 31]。测定数据按同一地区进行样地平均。
1.2.5 数据分析
按样地统计群落学特征和环境因素等数据,采用Microsoft Excel进行数据的初步计算及处理(平均数、标准差等);采用Sigmaplot软件进行气候条件和根系指标的图形处理;SPSS软件进行单因素方差分析(one-way ANOVA),并结合Duncan检验比较参数差异性;Canoco for Windows软件进行RDA分析(Redundancy Discriminate Analysis)。
2 结果与分析
2.1 辽东栎幼苗生物量沿环境梯度的变化
植物生物量的变化是植物自身与环境因素共同作用的结果,它既反映了植物对环境条件的适应及利用状态,也反映了植物本身的生存能力和生长发育规律[18]。随着年龄的增加,相同地区辽东栎幼苗地上、地下单株生物量逐渐增大(图2)。3个地区的辽东栎幼苗地上、地下单株生物量随年龄的增大有着显著的差异性变化(P<0.05),1年生幼苗地上、地下单株生物量太白地区高于黄龙和延安地区,随着辽东栎幼苗的发育,5年生幼苗地上、地下单株生物量黄龙地区高于延安和太白地区。辽东栎幼苗生物量受到生境变化制约的不同,不同年龄幼苗地上生物量存在一定的差异,而且这种差异随着年龄的增长有扩大的趋势。太白地区辽东栎幼苗1—5年根冠比依次为:0.79、0.77、0.79、0.71、0.66;黄龙地区辽东栎幼苗1—5年根冠比为:0.77、0.94、0.96、0.81、0.89;延安地区辽东栎幼苗1—5年根冠比为:0.80、0.86、0.81、0.75、0.77。
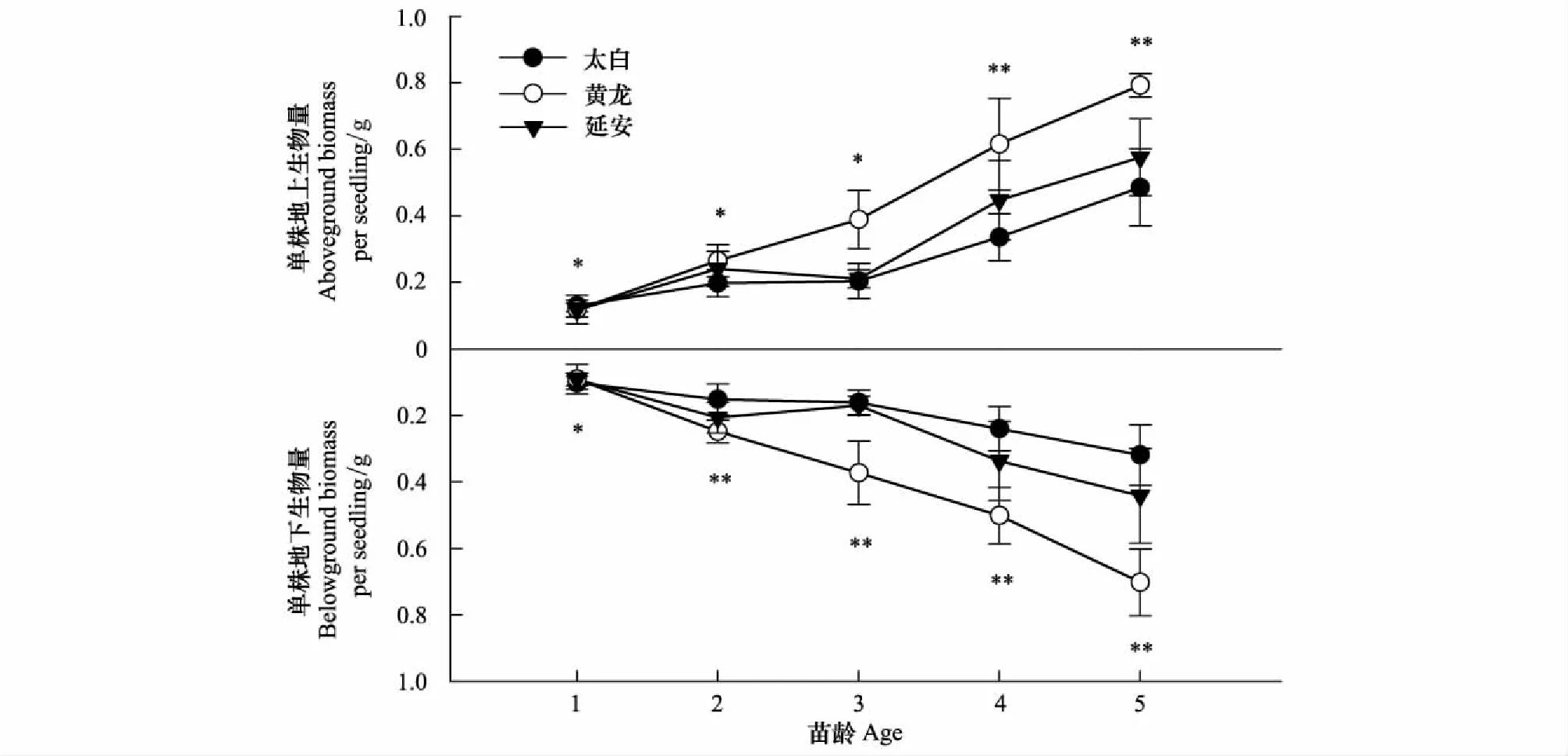
图2 不同地区辽东栎幼苗单株生物量
2.2 辽东栎幼苗根系形态沿环境梯度的变化
随着幼苗年龄的增加,相同地区的辽东栎1—5年生幼苗根系总长度、表面积、总体积、根尖数、分叉数、平均直径、比根长和组织密度均呈现增大趋势(图3)。不同地区辽东栎幼苗根系形态差异显著(P<0.05)。从秦岭北坡到黄土高原中部,随着幼苗的发育,根系总长度、表面积、总体积、根尖数、平均直径和组织密度最大值均出现在黄土高原南部黄龙地区。与黄龙地区相比,太白地区5年生幼苗根系总长度、表面积、总体积、根尖数、平均直径和组织密度分别下降了5.76%、37.33%、51.95%、10.31%、29.76%和5.39%;延安地区5年生幼苗根系总长度、表面积、总体积、根尖数、平均直径和组织密度分别下降了16.31%、26.64%、30.56%、19.21%、14.3%和9.31%。太白和黄龙地区,4—5年生辽东栎幼苗根系总长度、根尖数、分岔数和组织密度明显高于延安地区。降水相对较少的延安地区辽东栎幼苗根系在发育前期(1—2年)通过根系伸长、表面积增大和根尖数增多来适应干旱胁迫环境,随着幼苗的发育,受土壤水分限制,3年生以上幼苗的根系生长低于太白和黄龙两地。太白地区辽东栎幼苗根系通过减少体积,提高根系分岔数,促进根系生长发育;黄龙地区幼苗根系在土壤中所占体积较大,根系分岔数较小,根系主要以伸长生长来适应环境;延安地区的辽东栎幼苗通过增加根系比根长来适应半干旱胁迫环境(图2,图3)。

图3 不同地区辽东栎幼苗根系形态
太白、黄龙和延安辽东栎幼苗根系总长度、表面积、总体积、根尖数、分岔数、平均直径、比根长和组织密度随年龄的增加均呈线性函数(y=ax+b,a>0)变化格局(图4)。其中,幼苗根系总长度、表面积、总体积、根尖数和分岔数与幼苗年龄显著正相关(P<0.05),幼苗根系平均直径、比根长和组织密度与幼苗年龄没有显著关系(P>0.05)。
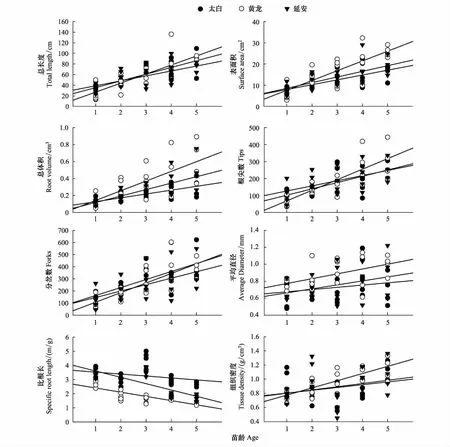
图4 不同地区辽东栎幼苗根系形态随年龄的变化
2.3 辽东栎幼苗根系分级伸展指标随年龄的变化
根据幼苗根系直径分布权重将不同地区辽东栎幼苗根系直径分为6个径级。径级范围依次为:0—0.2 mm,0.2—0.5 mm,0.5—1 mm,1—2 mm,2—4 mm和>4 mm,通过分级比较同龄幼苗长度、表面积和根尖数所占百分比。同一地区,各径级根系长度、表面积和根尖数所占百分比在不同年龄均有显著差异(P<0.05),根系长度和根尖数均随根系分级的增加而减少,表明辽东栎幼苗主要是通过低级根系的生长来适应环境,辽东栎幼苗均表现出较强的根系分支能力。与黄龙地区5年生幼苗相比,0.2—0.5 mm根系长度,太白地区下降了27.31%,延安地区下降了36.04%;0.2—0.5 mm根系表面积,太白地区下降了34.93%,延安地区下降了36.94%;0.2—0.5 mm根系根尖数,太白地区下降了46.73%,延安地区下降了49.53%(图5)。对不同地区辽东栎幼苗根系长度、表面积和根尖数占总数量的比例进行分析,发现各径级根系长度0—0.2 mm径级所占比例最大,太白、黄龙和延安0—0.2 mm径级所占总根系长度的比例分别为42.34%、32.32%和38.03%;各径级根系表面积占总数量的比例均是2—4 mm径级部分所占比例最大,太白、黄龙和延安2—4 mm径级所占总表面积的比例分别为32.62%、34.26%和31.12%;各径级根系根尖数0—0.2 mm径级所占比例最大,太白、黄龙和延安0—0.2 mm径级所占总根尖数的比例分别为82.78%、79.73%和82.53%。
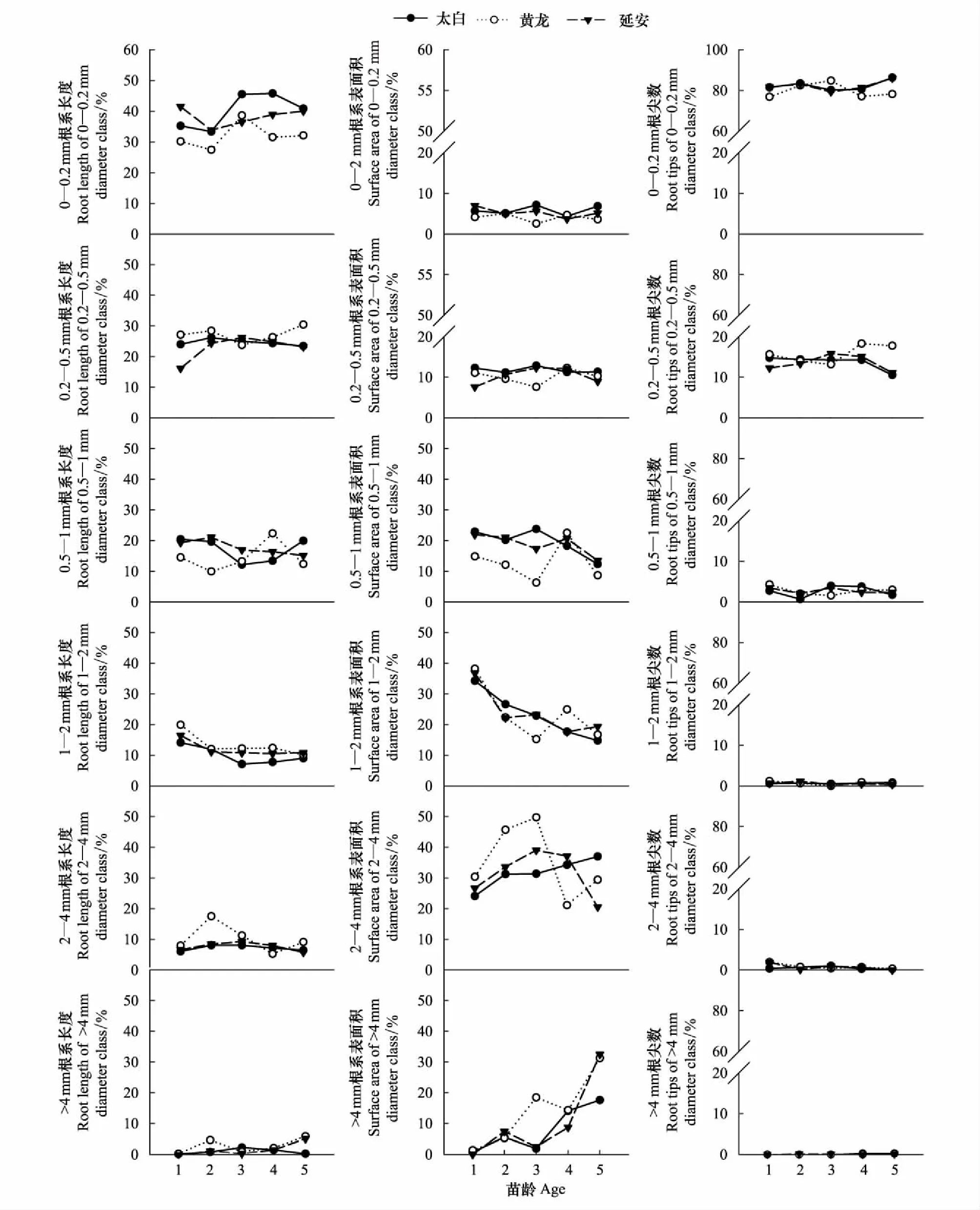
图5 不同地区辽东栎幼苗根系分级伸展指标所占百分比/%
2.4 辽东栎幼苗根系形态与环境因子的关系
对三个地区辽东栎幼苗根系指标与环境因子进行冗余分析(RDA),结果对变量的解释量达到14.7%,前两轴分别解释14.5%和0.2%(图6)。其中,对第1主分量贡献较高的环境因子包括速效磷(矢量值-0.5267)、硝态氮(-0.4914)、速效钾(-0.4777)、pH值(0.4662)、年均温(0.4524)、降雨量(-0.4118)、砾石含量(-0.4101)和速效氮(-0.4079);对第二主分量贡献较高的环境因子包括铵态氮(-0.2921)、有机质(-0.2833)、全氮(-0.2829)、夏季气温(0.2578)、速效氮(-0.2200)、砾石含量(-0.2191)和降雨量(-0.2183)。通过RDA双序轴第一轴可以看出,根系分岔数、总长度、比根长和根尖数与速效磷、硝态氮、速效钾、降雨量、石砾含量和速效氮的向量方向一致,具有正相关,与夏季气温、年均温和土壤pH值向量方向相反,具有负相关。
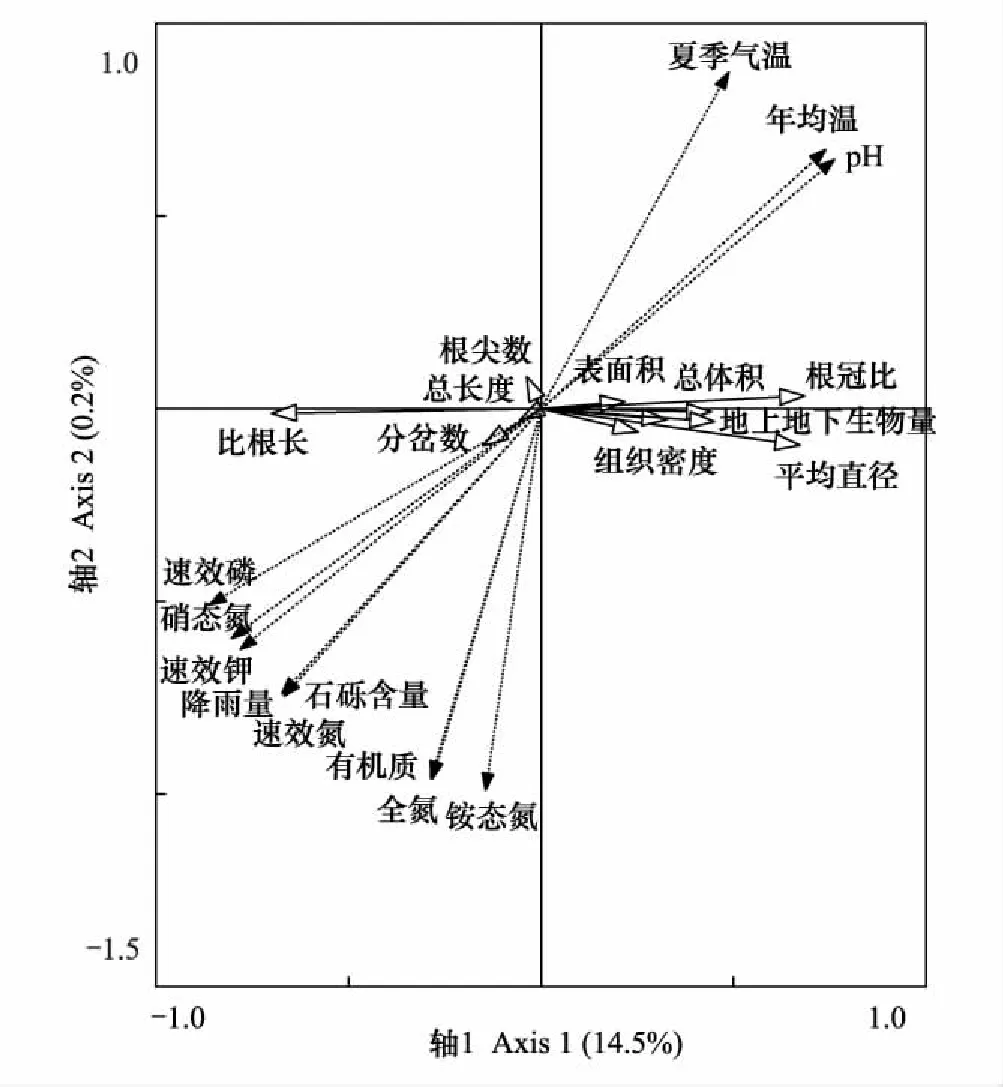
图6 辽东栎幼苗根系指标与环境因子冗余分析双序图
3 讨论
3.1 辽东栎幼苗根系形态沿环境变化的特点
植物的分布和生长受环境影响,严酷生境对植物器官发育有很强的抑制作用[32- 34]。当环境发生改变时,植物根系通过生物量积累和形态方面的改变来适应环境变化。与其他栎类相比,陕西地区辽东栎适合生长的年均气温为5—10℃,年均降水量为500—1000 mm[35]。适宜的水热条件,更有利于辽东栎幼苗的物质积累和能量分配。通过年龄研究幼苗根系形态结构,可以更好阐明根系对环境因子的利用效率。本研究发现,不同地区辽东栎幼苗均有较大的根冠比,幼苗早期地下生物量较小,为适应环境通过根系伸长生长来维持其对土壤养分水分的需求,经过一段时间的发育幼苗将更多的生物量分配给地下,增大根冠比以适应胁迫环境(图2)。随着辽东栎幼苗的发育,太白、黄龙和延安地区1—5年生幼苗根系均有较快生长,辽东栎幼苗根系总长度、表面积、总体积、根尖数和分岔数呈线性函数变化格局,可以用线性函数方程y=ax+b(a>0,P<0.05)进行描述(图4)。在半湿润气候的黄龙地区辽东栎幼苗根系前期(1—2年)生长低于太白、延安两地,随着幼苗发育,黄龙地区3年生以上的幼苗根系生物量和形态特征高于太白和延安地区,干旱胁迫下幼苗通过根系的伸长生长来适应胁迫环境,这与干旱胁迫下荒漠植物根系研究结果一致[36]。比根长反映根系生理活性的强弱,与土壤资源有效性密切相关。有研究表明,水曲柳和落叶松细根比根长介于1.81—117.74 m/g之间[37],榉树细根比根长在1.05—80.39 m/g之间[38]。本研究发现不同地区辽东栎1—5年生幼苗根系比根长介于1.28—4.46 m/g之间,且随着幼苗年龄增加而显著降低。幼苗根系比根长变化的拐点都出现在2—3年生之间,表明辽东栎幼苗是通过低级根来实现对土壤水分养分的有效利用。较高的组织密度表明根系对土壤养分水分具有较强的吸收能力(图3)。
3.2 辽东栎幼苗根系分级伸展指标随年龄变化的特点
植物根系系统中,小径级的根系末端具有水分和养分吸收功能。按照直径将根系划分成不同径级,不同径级根系对环境的敏感性存在差异[39]。本研究发现不同地区辽东栎幼苗根系长度主要集中在0—0.2 mm径级,占根系总长度的32%—42%。有研究表明,黄土高原地区1—3年生辽东栎幼苗2 mm以下的根系占总长度的92%[23];陕西栓皮栎分布区1—4年生栓皮栎幼苗0.5 mm以下的根系占总长度的55%—80%[40]。较细的根系有利于植物对水分和矿质元素的吸收,满足植物生长发育需求。不同地区辽东栎幼苗根系表面积主要集中在2—4 mm径级,占根系总表面积的25%—31%;根尖数主要集中在0—0.2 mm径级,占根系总根尖数的79%—83%(图5)。这表明在分布区内,辽东栎幼苗有足够的表面积和根尖数来进行物质循环[41- 42]。在降水较多、土壤水分养分条件较好的太白地区,根系主要是通过提高分岔数来拓展营养空间以适应环境;半湿润(黄龙)和半干旱(延安)的辽东栎幼苗主要是通过根系伸长生长来适应胁迫环境。
3.3 辽东栎幼苗根系形态与环境因子的关系分析
不同地区气候和土壤的差异能够解释辽东栎幼苗根系形态的差异。辽东栎幼苗根系分岔数、比根长、总长度、根尖数、石砾含量和降雨量呈较高的正相关,而增加土壤中速效磷、硝态氮和速效钾的含量能够提高幼苗比根长、分岔数、总长度和根尖数(图6)。说明辽东栎幼苗根系的生长和形态构成受环境的影响。有研究表明,黄土丘陵区植物根系长度受土壤有效磷、有效氮以及土壤水分的影响[43];在对栓皮栎幼苗根系生长与环境因子分析中,发现土壤氮含量、pH值和含水量对幼苗根系贡献率较高[40];铵态氮含量是影响油松幼苗根系生长的主要土壤因子[44];这与本研究结果一致。
从秦岭北坡到黄土高原中部,辽东栎幼苗根系表现出较好的形态可塑性。随着幼苗的发育,黄龙地区3年生以上幼苗根系生物量和形态特征高于太白和延安地区,幼苗通过根系的伸长生长来适应环境。在降水相对较多的太白地区,土壤水分通过降水得到有效补充,但因年均温度低生长年限短导致幼苗根系直径相对较小。在半干旱气候的延安地区,干旱胁迫导致根系分岔数少,根系表面积、总体积和平均直径高于太白地区。低级根(0—0.2 mm)通过增加根系长度和根尖数来获取水分和养分,高级根(2—4 mm)通过增加根系表面积来提高固定土壤能力。这是辽东栎幼苗为适应环境采用的一种获取资源的有效策略。土壤中一定的石砾含量可促进辽东栎幼苗根系生长,较低的降雨量会影响幼苗根系正常发育。在未来辽东栎林抚育经营中,通过合理抚育改善林地土壤质地,适当的控制表层土壤石砾含量,3%左右的石砾含量(>1 mm)和湿润土壤生境更有利于幼苗根系生长,达到提高林地更新能力的目的。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
广西林业科学(2021年6期)2022-01-21
辽宁省博物馆馆刊(2020年0期)2020-08-13
陶瓷科学与艺术(2019年10期)2019-12-18
东坡赤壁诗词(2019年3期)2019-07-05
娃娃乐园·综合智能(2018年17期)2018-10-09
当代陕西(2017年12期)2018-01-19
小学生作文(低年级适用)(2017年3期)2017-07-06
中国边疆民族研究(2016年0期)2016-12-18
