
厦门湾大型底栖动物分类学多样性指数及分类充分性
2018-09-19 08:26朱晓芬俞炜炜林俊辉黄雅琴廖建基
生态学报 2018年15期
朱晓芬,陈 彬,俞炜炜,林俊辉,黄雅琴,廖建基
国家海洋局第三海洋研究所,厦门 361000
《生物多样性公约》将生物多样性定义为“所有来源的生物体的变异性,包括陆域、海洋和其他生态系统和生态复合体,包括种内多样性、种间多样性和生态系统多样性[1]”。生物多样性的定义十分广泛,不仅包括物种的多样性,还涵盖分类的、遗传的、历史的和系统发育的多样性[2]。目前,生物多样性计算和评价通常采用物种数、Margalef丰富度指数、Shannon-Wiener多样性指数、Pielou均匀度指数等传统的多样性指数[3- 5]。然而,它们只是综合了一个群落内物种相对数量的信息量,将群落中每个物种视为等同,而未考虑物种间的差异及其亲缘关系[6]。同时,传统的多样性指数容易受到不同的样方大小、采样方法、生境类型或复杂程度的影响[7- 8]。基于此,1995年,Warwick 和Clarke[9]提出分类学多样性指数的测量方法,不仅考虑了丰度分布,还将物种间的分类学亲缘关系考虑在内。分类学多样性指数用来度量和解释群落中种类间形态关系的差异,它根据种类间分类关系的路径长度量化群落的分类多样性和差异性[10- 11],主要有分类多样性指数Δ、分类差异指数Δ*、平均分类差异指数Δ+和分类差异变异指数Λ+。
目前,尽管国际上分类学多样性指数在海洋大型底栖动物[12- 13]、自由生活线虫[14]及鱼类[15- 16]等生物群落得到一定应用,但是我国关于分类学多样性的研究主要集中于鱼类,而关于大型底栖动物的分类学多样性的研究非常有限,如曲方圆等[17]分析了黄海大型底栖生物的分类多样性;刘晓收等[18]研究了渤海大型底栖动物分类学多样性及其与环境因子的关系;胡成业等[13]对比了浙江6 个列岛潮间带大型底栖动物的分类学多样性。分类学多样性指数为现有的生物多样性方法补充一些有价值的信息[19],然而当前已有的这些研究多数仅计算了分类学多样性指数中的平均分类差异指数Δ+和分类差异变异指数Λ+两个指数,并且默认了该指数的适用性,并没有分析其与传统多样性指数的关系和差异,并缺乏对其适用性的深入探讨。
本文以厦门湾为研究区域,收集2014—2015年大型底栖动物的调查数据,计算厦门湾大型底栖动物的分类学多样性指数,分析了分类学多样性指数与传统生物多样性指数的相关性和依从性,探讨大型底栖动物的分类充分性,期望能为海湾生态环境的监测和评价提供新的思路和方法。
1 研究方法
1.1 研究区域与数据来源
研究区域厦门湾位于福建省南部、台湾海峡西南,为半封闭型海湾,包括九龙江口、西海域、东部海域、南部海域、同安湾、大嶝海域等。本研究收集了2014年5月和2015年5月航次的调查数据,共有32 个调查站位(图1),其中位于同安湾和大嶝海域的S17、S19—S21、S23、S25—S32等13个站位数据来源于2014年5月航次;位于同安湾的S18、以及西海域、九龙江口和南部海域的19 个站位数据来源于2015年5月航次。由于研究区域主体位于厦门珍稀濒危物种国家级自然保护区,2014—2015年在西海域、九龙江口和南部海域等区域均未实施大型海洋工程,而且底栖生物群落结构又是相对稳定的,故在随后的研究中将上述邻近两年相同月份的所有底栖生物调查数据一并进行统计分析。大型底栖动物的调查采用0.05 m2的抓斗式采泥器,每站连续采样5 次(合计采样面积为0.25 m2),所采泥样放入底栖生物漩涡分选器或套筛中冲洗,并用网目为0.5 mm的过筛器分选生物标本。生物样品置样品瓶中用5%福尔马林溶液固定保存后带回实验室。所有样品均鉴定到种。

图1 研究区域及大型底栖动物调查站位Fig.1 Study area location and station distribution
1.2 数据处理与分析
大型底栖动物物种名录通过WoRMS网站(http://marinespecies.org)查询和校对而构建。分类学多样性指数选取分类多样性指数(Taxonomic diversity, Δ)、分类差异指数(Taxonomic distinctness, Δ*)、平均分类差异指数(Average taxonomic distinctness, Δ+)和分类差异变异指数(Variation in taxonomic distinctness, Λ+)[20],各指数的含义和公式见表1。同时,选取物种数S、Shannon-Wiener多样性指数(H′)、Simpson多样性指数(D)、Margalef丰富度指数(d)、Pielou均匀度指数(J)等传统多样性指数。采用多元统计软件PRIMER 6.0计算物种水平的所有多样性指数,其中分类学多样性指数利用软件包中的TAXDTEST计算,各分类等级多样性权重值见表2。然后,利用SPSS 17.0软件中的Pearson Correlation Analysis计算传统多样性指数与分类学多样性指数的相关系数,定量分析两者之间的相关性。

表1 分类学多样性指数的含义及计算公式
Xi为第i个种类数量;ωij为第i个和j个种类在分类树中的路径长度;S为群落中出现的种类数

表2 各分类等级多样性权重值
为探讨大型底栖动物的分类充分性,在物种水平的基础上,分别将分类提高至属级和科级水平,统计不同分类水平的大型底栖动物丰度,利用PRIMER 6.0软件,采用与物种水平相同的方法计算属级、科级的所有多样性指数。首先,利用SPSS 17.0软件计算种级、属级和科级三个不同分类水平的同一多样性指数间的Pearson相关系数,并通过一元线性回归检验各指数间的关系,以分析不同分类水平的同一多样性指数之间的关系。然后,利用PRIMER 6.0软件,创建不同分类水平的“站位-底栖生物丰度”矩阵,对各站位数据进行开四次方根转换,分别计算各分类水平站位间的Bray-Curtis相似性系数,然后采用基于秩大小的非度量多维度标度(nMDS)方法分别绘制种级、属级、科级水平各站位的二维排序图,以对比分析不同分类水平所反映的底栖生物群落结构的一致性;应用2-STAGE(Second-Stage)方法计算不同分类水平矩阵之间的相似度,绘制聚类树,以分析不同分类水平群落之间的相似性。
2 结果
2.1 厦门湾分类多样性
厦门湾共采集到大型底栖动物247 种,隶属于11 门15 纲41 目110 科196 属,其中环节动物129 种,占总数的52.23%;节肢动物56 种,占总数的22.67%;软体动物34 种,占总数的13.77%;棘皮动物10 种,占总数的4.05%;刺胞动物和纽形动物各5 种,各占总数的2.02%;其他动物8 种,占总数的3.24%。
厦门湾大型底栖动物分类多样性指数(Δ)介于6.04—83.71之间,个别站位出现低值,站位差异较大,平均值为68.26;低值主要分布在大嶝海域的S27站(6.04)、S30站(28.95)及九龙江口的S5站(32.68),其余各站均高于50;高值主要分布在同安湾口的S21站(83.71)、东部海域的S24站(83.43)和西海域的S16站(81.54)。分类差异性指数(Δ*)介于74.27—99.54之间,各站的差异相对较小,平均值为84.23;低值主要分布在东部海域的S25站(75.26)、大嶝海域的S30站(75.39)和东部海域的S23站(76.11);高值主要分布在大嶝海域的S27站(99.54)、九龙江口的S5站(98.10)和同安湾的S17站(93.64)。
根据物种名录,计算得到厦门湾大型底栖动物平均分类差异指数(Δ+)和分类差异变异指数(Λ+)的理论平均值及95%置信漏斗曲线(图2),厦门湾平均分类差异指数(Δ+)介于76.01—89.12之间,理论平均值为86.82,S8、S10、S18、S27站位显著低于95%置信区间的下边界,低于95%置信区间通常意味着环境受到了扰动;分类差异变异指数Λ+介于276.42—498.46之间,理论平均值为345.0,S1、S18、S23、S25等多个站位高于95%置信区间上边界,这些站位分类差异变异指数略高,物种间的亲缘关系均匀程度较差。

图2 平均分类差异指数Δ+和分类差异变异指数Λ+的95%置信区间漏斗图Fig.2 95% probability funnels of average taxonomic distinctness and variation in taxonomic distinctnessS1—S32为32个调查站位
2.2 分类学多样性指数与传统多样性指数的相关性
通过计算大型底栖动物的传统多样性指数与分类学多样性指数的相关系数可知(表3),分类学多样性指数中的分类多样性指数Δ与Margalef丰富度指数、均匀度、Shannon-Wiener指数、Simpson指数之间均呈显著的正相关,相关系数分别为0.47(P<0.01)、0.96(P<0.01)、0.89(P<0.01)和0.97(P<0.01);分类差异指数Δ*与物种数、Margalef丰富度指数、均匀度、Shannon-Wiener指数、Simpson指数之间呈显著负相关,相关系数分别为-0.50(P<0.01)、-0.54(P<0.01)、-0.44(P<0.05)、-0.52(P<0.01)和-0.50(P<0.01),同时各指数间呈显著线性相关,拟合度较高。平均分类差异指数(Δ+)和分类差异变异指数(Λ+)与其他多样性指数间无显著相关性。
2.3 不同分类水平的同一生物多样性指数间的相关性
根据不同分类水平传统的多样性指数的相关系数计算结果(表4),不同分类水平的同一多样性指数之间(如物种数与属数、科数)表现出非常强的相关性,相关系数均大于0.95(P<0.01)(表4),并且呈线性相关,拟合度高(R2﹥0.9)(图3);与此同时,同一多样性指数的种级与属级的相关系数(0.99≤R2≤1.00,P<0.01)要明显高于种级与科级(0.96≤R2≤1.00,P<0.01)。类似地,不同分类水平的同一分类学多样性指数之间(如Δ(种)和Δ(属))也表现出非常强的相关性,相关系数大于0.70(P<0.01)(表4),且呈线性相关,拟合度高(多数R2﹥0.8)(图4);同一分类学多样性指数的种级与属级相关系数也明显高于种级与科级,如分类差异变异指数Λ+(种)与Λ+(属)的相关系数为0.90(P<0.01),而Λ+(种)与Λ+(科)的相关系数为0.70(P<0.01)。
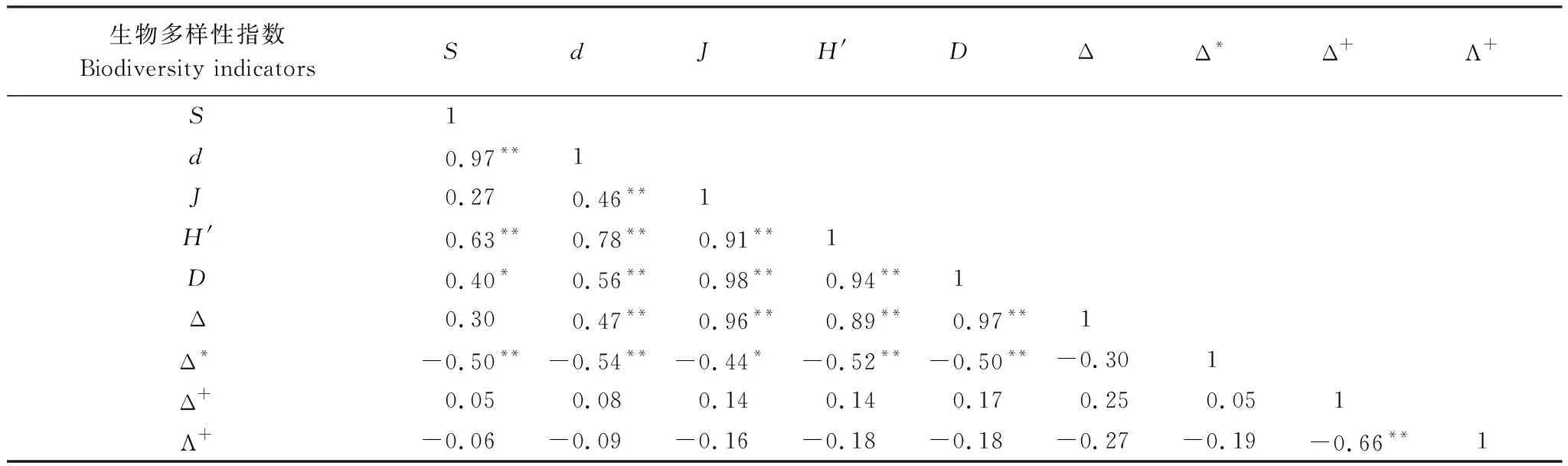
表3 生物多样性指数间的Pearson′s相关系数(种级)
**:P<0.01水平上显著相关;*:P<0.5水平上显著相关;S:物种数,Number of species;d:Margalef丰富度指数,Margalef richness index;J:Pielou均匀度指数,Pielou′s evenness index;H′:Shannon-Wiener多样性指数,Shannon-Wiener diversity index;D:Simpson多样性指数,Simpson diversity index

表4 各级分类水平间同一多样性指数的Pearson′s相关系数
**:P<0.01水平上显著相关;*:P<0.5水平上显著相关


图3 同一传统生物多样性指数在不同分类等级间的关系Fig.3 Relationship between the same traditional indicators of biodiversity at different taxonomic levelsH′:Shannon-Wiener多样性指数,Shannon-Wiener diversity index;d:Margalef丰富度指数,Margalef richness index;J:Pielou均匀度指数,Pielou′s evenness index;D:Simpson多样性指数,Simpson diversity index
2.4 种、属、科分类水平群落结构
根据32个站位不同分类水平的nMDS二维排序图(图5)可知,将鉴定水平从物种提高至属级时,除个别站位(如S4、S27),各站位之间的相对距离和位置变化非常小;将鉴定水平提高至科级时,各站位之间的相对距离和位置无明显的变化,但较物种至属级变化更为明显,如站位S5、S18、S24和S28。此外,拟合系数(stress)分别为0.23和0.24,介于0.10—0.25之间[21],拟合系数较低,表明数据结果可靠。nMDS结果表明,种级、属级和科级的群落结构一致性强,同时种级与属级的一致性较种级与科级更强。
根据2-STAGE(Second-Stage)方法得到不同分类水平底栖动物的聚类树状图(图6)可知,3种不同分类水平处理下底栖动物群落之间的相似性都较高(﹥80%),其中种与属的相似系数达92.35%,种与科的相似系数为80.17%。通过比较也可以看出,种与属的相似性较高,与种级相比,属级水平丢失的信息小于8%,而科级约20%,这与nMDS排序图结论相吻合。
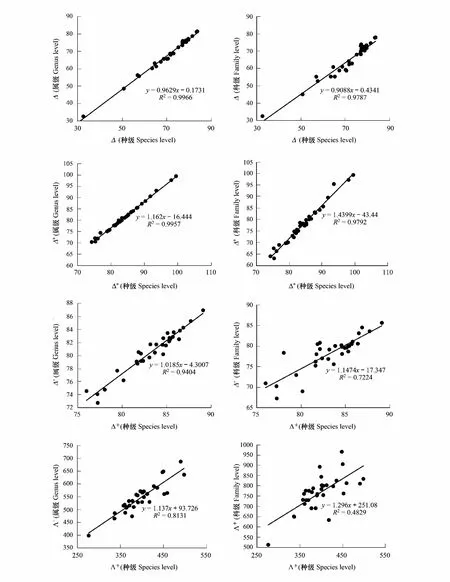
图4 同一分类学多样性指数在不同分类等级上的关系Fig.4 Relationship between the same Taxonomic distinctness index at different taxonomic levelsΔ:分类多样性指数,Taxonomic diversity;Δ*:分类差异指数,Taxonomic distinctness;Δ+:平均分类差异指数,Average taxonomic distinctness;Λ+:分类差异变异指数,Variation in taxonomic distinctness

图5 不同分类水平的nMDS二维排序图Fig.5 Non-metric multidimensional scaling ordinations at different taxonomic levels

图6 不同分类水平的聚类树状图 Fig.6 Dendrogram based on macrobenthos abundance at different taxonomic levelsa种级水平;b属级水平;c科级水平
3 讨论
3.1 厦门湾大型底栖动物的分类学多样性
厦门湾大型底栖动物分类多样性指数(Δ)介于6.04—83.71之间,均值为68.26,站位分布不均匀,最低值位于大嶝海域的S27站,仅有6.04。分类差异性指数(Δ*)介于74.27—99.54之间,均值为84.23,站位分布相对均匀,在S5站、S17站和S27站出现较高值。对比两指数可知,S5站和S27站出现Δ低而Δ*高的现象,这可能与样品中物种丰度不均匀有关,例如,S27站的菲律宾蛤仔(Ruditapesphilippinarum)丰度非常高,达4550 个/m2,占总丰度的97%。
根据平均分类差异指数(Δ+)的95%置信区间漏斗图,厦门湾大型底栖动物Δ+理论平均值为86.82,大部分站位Δ+分布在95%置信区间内,S8、S10、S18和S27等个别站位落在置信区间外,表明厦门湾局部区域环境受到了一定程度的干扰。根据分类差异变异指数(Λ+)的95%置信区间漏斗图,厦门湾大型底栖动物Λ+的理论平均值为345.0;S1、S18、S23、S25站位的Λ+值高于95%置信区间上边界,表明这些站位物种间的亲缘关系均匀程度较差。将这两个指数的计算结果相结合,S18站位的Δ+低于95%置信区间且Λ+高于95%置信区间,这表明该站位的环境受到了干扰。究其原因,S18位于同安湾,离岸很近,因此受到陆源污染及人类活动影响较大。
3.2 分类学多样性指数与传统多样性指数
从计算公式上看,分类多样性指数(Δ)和分类差异性指数(Δ*)是基于概率论且与优势度相关的指数,是Simpson指数的自然延伸,在Simpson指数的基础上,增加了不同种类在分类学差异方面的信息,既考虑了群落中种类的数量(丰度或生物量)和均匀程度,又将物种的分类地位考虑在内[22],因此与传统的多样性指数间存在显著相关性。与Δ相似,Δ*是对Δ简化后的指数,即在Δ的基础上忽略相同物种个体的分类路径长度[23]。这两个指数是综合性较强的指数,混合了分类关系、丰度及其分布均匀性信息,因此,分类亲缘关系、丰度及其分布的均匀性都可能影响这两指数的高低。本研究中,厦门湾S5站和S27站出现Δ低值,这可能很大程度上与物种丰度分布的均匀性相关,S27站的菲律宾蛤仔(Ruditapesphilippinarum)丰度特别高,达4550 个/m2,占总丰度的97%;类似地,S5站的光滑河兰蛤(Potamocorbulalaevis)丰度出现高值,达765 个/m2,占总丰度的81%。若将S5站和S27站中的丰度出现极值的物种剔除的话,S5站的Δ值由32.68升高至72.22,S27站的Δ值由6.04升高至60.23,d、J、H′、D等传统多样性指数也相应大幅度提高(表5)。

表5 S5和S27站位除去丰度极高种前后各多样性指数的对比
平均分类差异指数(Δ+)和分类差异变异指数(Λ+)在仅存在物种名录的情况下可以计算,计算得到的是一理论平均值,只考虑种类是否出现,而且不考虑物种的丰度数量[24- 25],从而理论上与传统的多样性指数相关性不强,这在本文厦门湾的研究中也得到验证。本研究中,厦门湾S5站和S27站中个别物种的丰度出现峰值,若将出现峰值的物种剔除的话,Δ+和Λ+的变化不显著,表明其与物种的丰度数量相关性不强(表5)。在4个分类学多样性指数中,Δ+和Λ+较Δ和Δ*运用更广泛,这两指数的优势可能在于:①数据要求低,在仅有物种名录存在的情况就可以计算,可适用于一些数据较欠缺的区域[26];②不依赖于取样大小和取样方法,可用于不同采样努力、不同区域、不同生境类型的比较;③通过漏斗图可快速识别受干扰的站位或区域,而不需要背景或参考值,解决了生态评价中确定参考值的困难。
生物多样性是一个多要素的概念,一般认为应包括3个方面,即物种的数量(丰富性)、各种类丰度的分布情况(均匀性)和不同种类之间的分类学关系(分类差异性)[27]。不同多样性指数从不同的角度反映多样性的信息,单一指数往往难以全面反映生物多样性的所有信息。与传统多样性指数相比,分类学多样性指数考虑了物种在分类以及功能上存在的差异。传统多样性指数将群落中每个物种同等对待,一个物种的减少可能对物种数的影响并不大,但是对分类学多样性指数的影响可能非常大,尤其是当一个门下只有一个种时,该种的消失就等于一个门的消失。例如,厦门湾S29和S31站位的大型底栖动物物种数都为23,多数多样性指数均表现出S31站略高于S29站;与传统多样性指数相比,这两站的分类学多样性指数的差异更为显著,变化也更为灵敏[25](表6)。这表明,在传统多样性指数难以区分两个站位时,利用分类学多样性指数就能够区分,这也反过来证明,物种丰富度和均匀性相似的两个站位,它们的物种间的亲缘关系可能存在很大差异。因此,较传统多样性指数而言,分类学多样性指数增加了分类学信息,可作为传统多样性指数的补充。

表6 S29和S31站位间多样性指数的对比
3.3 大型底栖生物的分类充分性
海洋生物分类是海洋生物多样性评价的基础。海洋生物分类是否准确会直接影响生物多样性评价的结果,尤其对于分类多样性指数的计算更为重要。1985年,Ellis[28]提出了“分类充分性”(Taxonomic Sufficiency,TS),并将其定义为:在任何项目中有机体必须被鉴定到一个能满足精确指示有机体生物学需要的水平。“分类充分性”在水生生态系统中已得到广泛运用[29- 32],用于探讨满足需求的生物鉴定水平。
在本文厦门湾大型底栖动物研究中,一方面,种级、属级、科级三个分类水平间的同一多样性指数(包括分类学多样性指数和传统多样性指数)表现出显著正相关,线性拟合度较强,且同一指数种级与属级的相关系数要明显高于种级与科级;另一方面,nMDS表明种级、属级和科级的群落结构一致性强,且种级与属级的一致性较种级与科级更强;2-STAGE的相似性和聚类结果也表明属级丢失的信息较科级少,种级与属级的相似性更高,群落结构更为一致,这与nMDS的结果一致。根据以上的结果可看出,从属级或科级评价厦门湾大型底栖动物的多样性与种级的结果大体是一致的,但是与种相比,科和属所反映的信息更少,并且科所丢失的信息(20%)要比属(8%)显著增多。这结果与一些已有的研究相一致,例如,吴浩东等[33]在研究大型无脊椎动物不同分类阶元对水生态评价的影响中发现,与种级相比,属级水平丢失的信息小于5%,科级为7%。
不少研究表明[34-36],将生物鉴定至属、科甚至更高的分类水平足以满足需求,甚至还有研究认为科、属水平的评价结果比种水平更为准确[37]。Gray等[38]指出物种数据中有很大程度的信息冗余,将数据聚合到更高的分类水平群组一定程度上可弱化这个问题。Warwick[39]也指出自然变异主要通过物种更替影响群落结构,污染效应从更高的分类水平上也能监测到,种水平的鉴定在污染评价中经常是不必要的。
在海洋调查中,我国学者通常将海洋生物鉴定至物种水平,但这却也一直备受争议。海洋物种水平的鉴定存在诸多现实的问题,如鉴定成本高、工作量大、准确性受分类人员的鉴定水平的影响大[40]。将海洋生物鉴定至科、属等更高的水平可减少调查费用[41],例如,Ferraro和Cole[42]估算出属、科、目和门水平的鉴定相对于种水平的鉴定成本分别减少了23%、55%、80%和95%。物种水平的鉴定难度较大,而不少监测机构都存在缺乏专业的生物鉴定人才的问题,更高水平的分类难度较小、可操作性较强,同时也减小了分类错误的风险。此外,对于临时突发事件,往往没有足够的资源和时间进行具体细致的分类调查[43- 44],较高水平的分类可以提供快捷的途径,为管理决策者作出有根据迅速的判断[45]。因此,在条件有限的情况下,可根据实际的需求适当放宽海洋生物的鉴定水平。
大型底栖动物分类充分性的理论研究与实际应用对环境监测调查与评价来说是很有意义的,本研究以厦门湾作为区域验证,其结果也支持这一观点。但是,由于生物鉴定到较高水平会导致一些生态信息的丢失[30][46],“分类充分性”的使用也存在争议。在某些情况下,例如在科学研究中,或对于一些数据较为缺乏的区域,有必要将生物鉴定到最低的物种水平,以获取更多的信息,进而展开深入透彻的研究和分析。因此,对于一些需要获取较全的信息才能满足科学研究、生态评价和管理等需求时,将生物鉴定至物种水平是有必要的。综上,生物鉴定水平的选取应该密切结合实际的需求,既要考虑避免大量人力、物力的浪费,同时也要考虑科学性和严谨性。
猜你喜欢
课堂内外·教师版(2022年4期)2022-05-23
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
人大建设(2019年5期)2019-10-08
人大建设(2019年3期)2019-07-13
党的生活(黑龙江)(2018年2期)2018-03-10
党的生活(黑龙江)(2017年7期)2017-07-19
中央民族大学学报(自然科学版)(2016年2期)2016-06-27
新疆大学学报(哲学社会科学版)(2015年6期)2015-10-12
中央民族大学学报(自然科学版)(2015年3期)2015-06-11
