
改性魔芋葡甘露聚糖对齐口裂腹鱼脂质代谢及相关基因表达的影响
2018-09-18 02:56:54吕珍珍邬应龙严秋萍陈明睿冯莉梅赵嘉琪王冰莹
食品与生物技术学报 2018年7期
吕珍珍, 邬应龙, 何 梅, 严秋萍, 陈明睿, 冯莉梅, 赵嘉琪, 王冰莹
(四川农业大学 食品学院,四川 雅安 625014)
葡甘露聚糖广泛存在于魔芋中,是一种不可消化的可溶性膳食纤维,但魔芋葡甘露聚糖在水中粘度大、溶解度低、流动性和溶胶稳定性差,使其应用受到限制。目前主要采用酶法、物理和酶酸结合的方法对葡甘露聚糖进行降解,提高其在水中的溶解度,扩大葡甘露聚糖的应用范围[1]。
齐口裂腹鱼是中国特有的重要冷水性经济鱼类,肉质鲜嫩,营养价值高。鱼类营养性脂肪肝及腹腔脂肪过度蓄积的情况极易发生,这不仅会妨碍鱼类的生长,甚至还可能引发鱼类疾病,增加养殖成本[2]。对齐口裂腹鱼脂质代谢方面的研究,主要集中在养殖中摄食蛋白质及脂肪含量等对鱼肉品质及脂质代谢影响,有关不同改性魔芋葡甘露聚糖对齐口裂腹鱼脂质代谢影响研究较少。
作者分别参考张辽[3]、高松[4]方法制备较低相对分子质量的酸解氧化魔芋和魔芋葡甘露聚糖硫酸酯,探讨在饲料中添加不同剂量两种不同改性方式的魔芋葡甘露聚糖对齐口裂腹鱼生长、血液、基因等脂质代谢方面的影响。
1 材料与方法
1.1 材料与试剂
氧化魔芋葡甘露聚糖(OKGM):四川农业大学食品学院功能性食品实验室提供。
高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)、总胆固醇(T-CHO)、甘油三酯(TG)试剂盒:南京建成生物工程研究所产品;琼脂糖:Sigma公司产品;Golden view 染料、2×Taq PCR Master Mix、DEPC-H2O、Loading Buffer、 DL 2000 Marker:天根(Tian gen)试剂公司产品;SYBR 荧光定量试剂盒、cDNA反转录试剂盒均、RNA提取Trizol:Vazyme试剂公司产品。
1.2 主要仪器
Bio-Rad实时荧光定量PCR仪、Bio-Rad C1000普通 PCR仪、Bio-Rad电泳仪、Varioskan Flash全波长扫描式多功能读数仪 (多功能酶标仪)、微量移液枪、ThermoFisher高速低温离心机等。
1.3 实验动物
从雅安市天全县鱼泉乡购得420尾平均鱼体质量为(80±1.21)g齐口裂腹鱼,购买后于作者所在实验室驯养15 d。
1.4 试验设计与饲养管理
驯养结束后,按试验设计将实验动物分为7个组(每组3个重复,每个重复20尾),分别为对照组(基础饲料)、质量分数 0.4%OKGMS、0.8%OKGMS、1.6%OKGMS、0.4%AOKGM、0.8%AOKGM 和 1.6%AOKGM。每天按每缸体质量2%投喂饲料,每日3次;早、晚对鱼缸进行换水,并清除缸内不溶物。饲养期60 d(12 h白昼,12 h夜晚),24 h不间断充氧,保持微流水。
1.5 生长性能指标测定
饲养60 d后,鱼体饥饿24 h,从每缸中选取5尾鱼称量鱼体质量,解剖后取出肝胰脏、背肌及肠道脂肪的质量,用预冷的无菌生理盐水冲洗干净,滤纸吸干水分,称质量,置于-20℃冰箱,备用。测定鱼的肥满度,肝体比和肠脂比。
1.6 肌肉、肝脏基础成分的测定
肝脏基础成分测定参照食品安全国家标准GB 5009.3-2010 《食品中水分的测定》105℃直接干燥法、GB/T 5009.6-2003《食品中脂肪的测定》索氏抽提法、GB 5009.5-2010《食品中蛋白质的测定》凯氏定氮法进行测定。
1.7 样品的采集和制备
称重后,尾静脉取血。将每尾鱼的血液,4℃条件下静置30 min待血液凝固后,于4℃条件下以3 000 g/min离心15 min,分离血清以用于血清中高密度脂蛋白胆固醇、低密度脂蛋白胆固醇、总胆固醇、及甘油三酯的测定。测定方法参照试剂盒说明书方法进行。
每缸另取5尾鱼取血后,解剖鱼体,分别迅速取出肝胰脏,迅速置于液氮中冷冻,再转入-80℃冰箱中保存,用于RNA提取。
1.8 肝脏脂质代谢相关基因定量分析
1.8.1 RNA提取 肝脏中RNA提取及纯化参照TaKaRa试剂盒方法进行。
1.8.2 后续实验为RT-PCR的cDNA合成 参照HiScript®1st Strand cDNA Synthesis Kit试剂盒说明书进行操作。
将产物cDNA稀释10倍保存于-20℃冰箱,用于后续荧光定量PCR反应。
1.8.3 脂质代谢相关引物 参照郑俏然[4]等设计的齐口裂腹鱼脂质代谢相关引物 β-actin,fas,lpl,hl,选用β-actin作为齐口裂腹鱼内参基因 (引物设计见表 1)。

表1 实时定量PCR引物序列Table 1 Real-time quantitative PCR primer sequence
1.8.4 荧光定量PCR 参考Tian等[5]已报道的方法稍作修改,每个样品3个平行。采用SYBR Green I荧光染料法,对 β-actin,fas,lpl,hl引物进行荧光定量分析:10 μL反应体系,含 5 μL SYBR® Premix Ex TaqTM (2×)、cDNA 1 μL、 上下游引物各 0.25 μL、DEPC 水 3.50 μL。 反应条件:95 ℃ 3 min;95 ℃10 s,相应退火温度 30 s,39个循环后,进行95℃处理10 s;扩增完毕后,迅速降温到65℃进行溶解曲线分析,然后以0.5℃/s的速率从65℃升温到95℃,连续测定样品荧光强度以获取溶解曲线。
参考Livak等[6]数据分析方法:比较对照组和基础组样本数据,采用2-ΔΔCT相关定量进行计算。
1.9 数据处理
采用SPSS22.0软件进行单因素方差分析,多重比较采用 Duncan’s检验法(P<0.05)。
2 结果与分析
2.1 OKGMS及AOKGM对齐口裂腹鱼基础生长指标的影响
与基础相比,OKGMS及AOKGM各添加剂量对鱼体体重、肥满度均无显著影响,当添加质量分数0.4%AOKGM时,显著增加鱼体的肝体比与肠脂比。
2.2 OKGMS及AOKGM对齐口裂腹鱼肝脏基础成分及血清生化指标的影响
与基础组相比,在OKGMS改性条件下,肝脏脂肪质量分数均有所降低,其中质量分数1.6%OKGMS差异显著;血清T-CHO质量分数均显著降低;血清HDL-C、TG含量均有所增加,其中0.4%OKGMS及0.8%OKGMS差异显著。与基础组相比,在AOKGM改性条件下,肝脏水分质量分数有所降低,其中0.4%AOKGM及0.8%AOKGM差异显著;肝脏脂肪、血清T-CHO均显著降低;0.8%AOKGM及1.6%AOKGM显著增加血清HDL-C及显著降低血清TG质量分数。
2.3 OKGMS及AOKGM对肝脏相关基因q-PCR的影响
2.3.1 齐口裂腹鱼lpl,hl,fasPCR扩增结果 以反转录肝脏cDNA为模板,进行PCR扩增,产物通过1 g/dL琼脂糖凝胶检测后发现,分别得到fas145 bp,lpl166 bp,hl268 bp目的产物,且条带清晰明亮,满足PCR分析要求。

图1 齐口裂腹鱼肝脏fas,lpl,hl pcr扩增结果Fig.1 Amplification results of fas,lpl,hl of Schizothorax prenanti
2.3.2 OKGMS 及 AOKGM 对肝脏lpl,hl,fas荧光定量的影响lplmRNA在肝脏中相对表达量见图2。与基础组相比,不同剂量OKGMS、质量分数0.4%AOKGM及1.6%AOKGM组肝脏中lplmRNA表达显著增加,0.8%AOKGM组增加2.7倍。
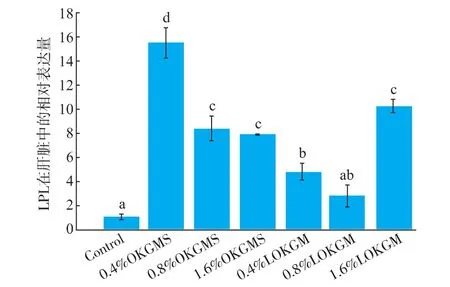
图2 lpl mRNA在肝脏中荧光定量PCR相对表达Fig.2 lpl mRNA Real-time quantitative PCR relative expression in the liver
hlmRNA在肝脏中相对表达量见图3。与基础组相比,不同剂量OKGMS及AOKGM,hlmRNA相对表达量没有显著差异。

图3 hl mRNA在肝脏中荧光定量PCR相对表达Fig.3 hl mRNA Real-time quantitative PCR relative expression in the liver
fasmRNA在肝脏中相对表达量见图4。与基础组相比,1.6%AOKGM显著增加了肝脏中fasmRNA相对表达量,其他各组没有显著差异。
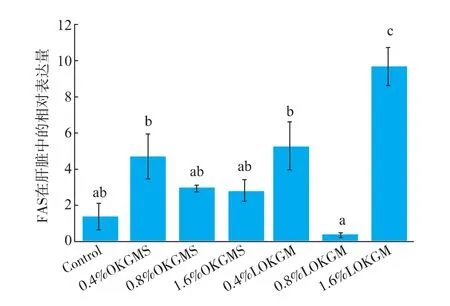
图4 fas mRNA在肝脏中荧光定量PCR相对表达Fig.4 fas mRNA Real-time quantitative PCR relative expression in the liver
3 讨 论
肝脏是鱼体脂类代谢的重要器官[7],在鱼体脂类消化、吸收、合成、分解等过程中起重要作用。
在陆地生物中发现,供给甘露寡糖(MOS)影响胆汁酸排泄、食糜上清液粘度或微生物方面变化影响肠脂质吸收,进而影响游离脂肪酸进入肝脏进行消耗[8]。然而在鱼体方面,膳食纤维对脂质消化吸收的机制还不太清楚。肥满度、肝体比、肠脂比反应鱼体脂肪含量,过多脂肪在鱼肉、肝胰脏及肠系膜等处积累,不仅造成鱼肉品质差,鱼体利用率下降影响鱼体品质,且易形成脂肪肝造成病害。添加一定剂量OKGMS及AOKGM不会影响齐口裂腹鱼生长效果,质量分数0.4%AOKGM显著增加齐口裂腹鱼肝体比、肠脂比。这与Mansour等[9]证实巨鲟鱼摄食质量分数0.2%active MOS 46 d脂质质量分数增加,而摄食质量分数0.3%Immunoster 8周脂质不变结果一致。然而Ye JD等[10]研究证实牙鲆鱼摄食质量分数0.5%Bio-MOS 56 d,降低了脂质沉积。这表明膳食纤维种类、饲喂时间、添加剂量等可能影响鱼体脂质消化与生长。
在鱼类脂肪合成和沉积过程中,lpl,hl,fas等发挥着重要作用。lpl和hl是调节机体脂肪沉积和脂质代谢的关键因子,fas是脂肪合成的关键酶,所以研究lpl,hl,fas基因表达量对研究鱼类脂肪代谢至关重要[11]。脂肪合成分为两个部分,第一部分:乙酰辅酶-A羧化酶通过碳酸氢盐和ATP催化羧化乙酰辅酶-A形成丙二酰辅酶-A;第二部分:脂肪酸合成酶在NADPH条件下催化缩合乙酰辅酶-A和C2结构形成丙酰辅酶A,然后形成棕榈酸[12]。fas在脂肪酸的从头合成中发挥重要作用,它能催化乙酰辅-A和丙二酸单酰辅酶-A合成长链饱和脂肪酸,对动物脂肪沉积有着重要意义。OKGMS及AOKGM增加了肝脏中lplmRNA表达,当添加质量分数1.6%AOKGM fas表达显著增加。Yokota等[13]证实了岩藻多糖对lpl基因表达的影响,其结果指出岩藻多糖能诱导脂肪细胞lpl基因表达,且其表达量随着岩藻多糖剂量增多及作用时间延长而增加。目前关于多糖对水生动物hl,fas基因表达的影响研究还很少。夏晓杰等[7]证实大中小鱼中lpl与fas表达成正相关关系,对肌间脂肪沉积有积极作用而hl作用不大。这表明OKGMS及AOKGM调控了lpl表达,在一定范围内影响fas表达,且随着膳食纤维种类及添加剂量不同表达不同。
脂肪在鱼类细胞内主要以甘油三酯形式存在,甘油三酯是鱼类脂类代谢的主要产物[14]。血浆中甘油三酯的含量是通过极低密度脂蛋白和乳糜微粒颗粒的合成与分解来调控。血液循环中是通过lpl和hl作用以及利用胆固醇酯酶转移蛋白使内脂蛋白与甘油三酯进行置换来清除富含甘油三酯的脂蛋白[15]。OKGMS能显著降低肝脏脂肪及血清胆固醇含量,增加血清高密度脂蛋白含量,当添加质量分数为1.6%OKGMS效果明显;AOKGM显著降低肝脏水分、脂肪及血清胆固醇、甘油三酯含量,增加血清高密度脂蛋白含量,当添加剂量为0.8%AOKGM效果明显。与Fiordaliso等[15]研究低聚木糖和低聚果糖减少了肝脏脂肪生成,血液和肝脏胆固醇和甘油三酯水平,增加了血液中HDL/LDL比率结果一致。Lahoz等[16]证实在哺乳动物中hl作为高密度脂蛋白配体是一种脂肪分解酶,促进肝脏对它吸收。而且鱼体中lpl和hl促进了肝脏对脂蛋白吸收,所以鱼血清中高密度脂蛋白水平比其他脊椎动物高几倍。Tian[5]等证实lpl表达增加,血清中高密度脂蛋白含量增加,甘油三酯含量降低。Delzenne等[17]证实较低肝脏脂肪可能与摄食中低聚木糖通过鱼肠道微生物菌群发酵,生成葡萄糖激酶与结肠中短链脂肪酸。因此Campbell等[18]证实老鼠饮食中含有低聚木糖增加了盲肠中短链脂肪酸即醋酸盐、丙酸盐、丁酸盐和乳酸盐浓度。然而主要短链脂肪酸产物醋酸盐和丙酸盐对脂质代谢调制是相反的。醋酸盐是脂肪生成的基质,丙酸盐竞争性抑制醋酸盐进入肝脏细胞。益生元被肠道微生物发酵的模式和肝脏醋酸盐与丙酸盐比例将决定其低脂肪性能,同时醋酸盐与丙酸盐比例因鱼种类的不同而改变。由此推测添加一定剂量OKGMS或AOKGM,可能会增加食物体积,稀释肠道内容物,促进有益菌乳酸菌、双歧杆菌生长,产生短链脂肪酸并提高肠道短链脂肪酸浓度及比例,调控齐口裂腹鱼肝脏中lpl表达,影响肝脏脂蛋白酯酶的活性,增加高密度脂蛋白含量促进高密度脂蛋白反向运输胆固醇进入肝脏,加速胆固醇转运与代谢,降低肝脏脂肪、血液中胆固醇和甘油三酯含量,增加高密度脂蛋白含量,调节鱼体脂质代谢。
4 结 语
对比了两种改性魔芋葡甘露聚糖对齐口裂腹鱼脂质代谢及肝脏相关基因的表达,发现在OKGMS改性条件下,不同添加剂量降低齐口裂腹鱼肝脏脂肪及血清胆固醇含量,显著增加血清高密脂蛋白含量及肝脏lpl表达,且添加质量分数为1.6%效果更好。在AOKGM改性条件下不同添加剂量降低肝脏脂肪;对血清胆固醇、甘油三脂含量呈先降低后增加趋势;增加血清高密脂蛋白含量及肝脏lpl表达,且添加质量分数为0.8%效果更好。
猜你喜欢
大理大学学报(2023年12期)2024-01-12 02:54:28
农业工程学报(2022年11期)2022-08-22 14:06:38
淡水渔业(2022年2期)2022-03-24 02:15:14
水电与抽水蓄能(2021年2期)2021-05-14 01:42:26
农业工程学报(2021年4期)2021-05-09 08:23:58
农业工程学报(2020年6期)2020-05-19 07:46:38
现代检验医学杂志(2016年1期)2016-11-12 13:19:54
医学研究杂志(2015年12期)2015-06-10 06:57:46
国际心血管病杂志(2015年5期)2015-02-27 12:11:35
中华皮肤科杂志(2014年3期)2014-12-19 12:54:57
