
奥斯特亚科(圆线目:毛圆科)线虫系统分类学研究概况
2018-09-15 01:55刘振斌杨学财万雪梅杜江龙蔡葵蒸
东北农业大学学报 2018年8期
刘振斌,杨学财,万雪梅,杜江龙,蔡葵蒸*
(1.西北民族大学甘肃省动物细胞工程技术研究中心,兰州 730030;2.西北民族大学生命科学与工程学院,兰州 730030)
毛圆科(Trichostrongylidae)线虫是野生和家养反刍动物常见寄生虫,深入了解其系统分类学对阐明寄生虫行为学(包括病原学)、流行病学、宿主与寄生虫间进化及生物地理历史等十分必要[1]。过去几十年,毛圆科系统分类学研究存在争议,Chilton等根据rDNA序列数据作进化分析,认为毛圆科由6个亚科组成,分别是Libyostrongylinae、纵纹亚科(Graphidiinae)、古柏亚科(Cooperiinae)、奥斯特亚科(Ostertagiinae)、毛圆亚科(Trichostrongylidae)和血矛亚科(Haemonchinae)[2]。其中奥斯特亚科是毛圆科重要成员之一,主要寄生于反刍动物第四胃[3]。传统上该亚科线虫根据雄虫形态学特征开展种的鉴定,但由于雄虫存在多态性、隐藏种及种复合体等现象,传统分类学方法面临挑战。鉴此,本文根据国内外研究现状,对奥斯特亚科系统分类学展开讨论并提出建议。
1 奥斯特亚科形态学特征
1.1 虫体特征
奥斯特亚科线虫头端无头泡,口囊退化,有微小口腔,颈乳突发达,雄虫交合伞发育良好,伞前乳突发达,交合刺较短,雌虫阴门位于虫体后半部,体表有若干条垂直于表面纵嵴,寄生于反刍动物。仅淡红猪圆线虫(Hyostrongylusrubidus)未见于反刍动物,分类地位尚无定论[4]。除经典著作描述该亚科线虫形态结构外,其体表纵嵴分布模式是重要发现[5],Lichtenfels等研究奥斯特亚科属体表纵嵴分布模式(见图1),认为不同属体表纵嵴分布不同,可根据颈乳突与食道后端体表区域纵嵴分布模式和体中部纵嵴数目分类[6-7]。
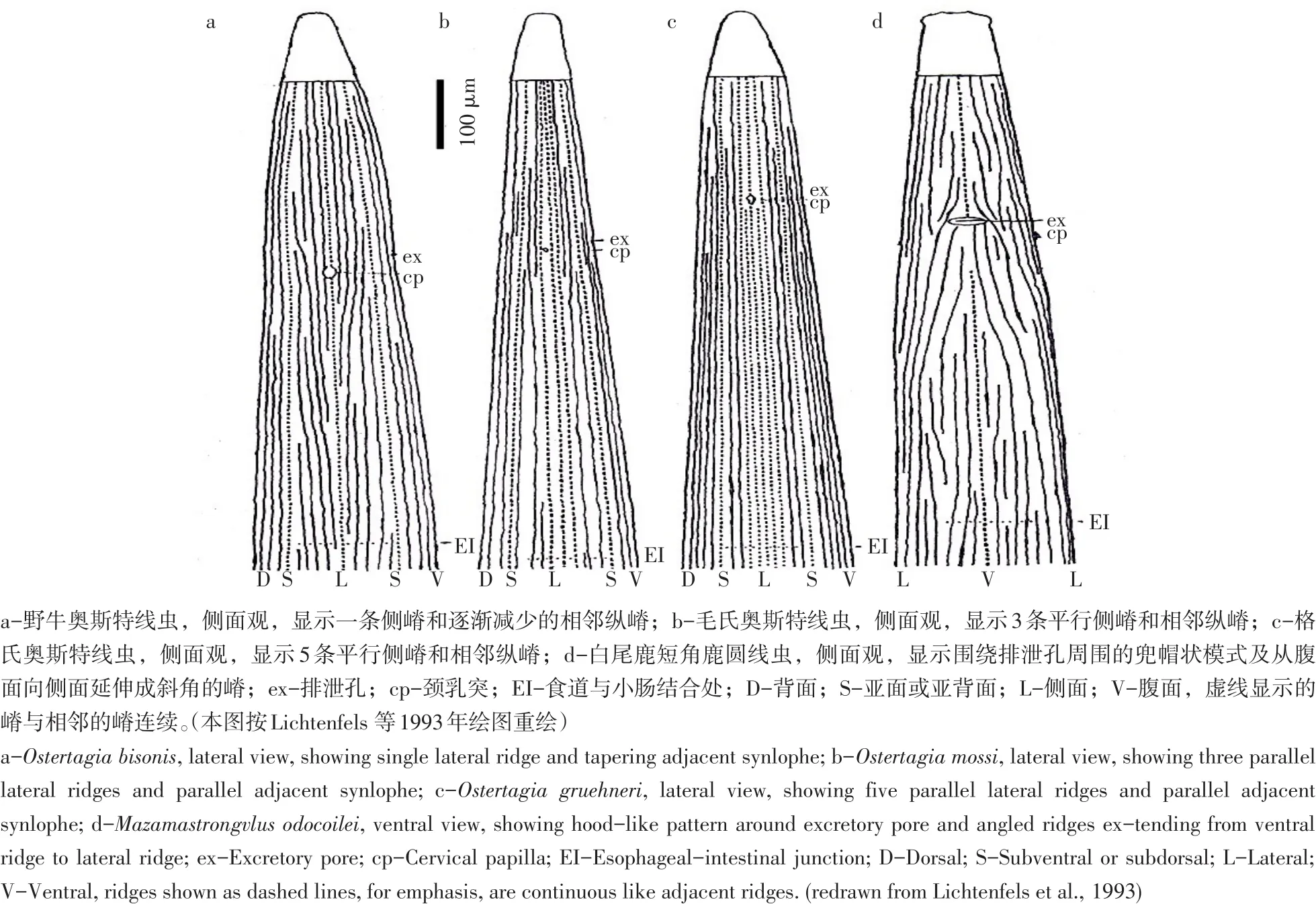
图1 奥斯特亚科食道区域体表纵嵴模式Fig.1 Synlophe patterns in the region of the esophagusof Ostertagiinae
1.2 雄虫尾端特征
奥斯特亚科交合伞侧肋排列模式分两类[8-9],一类以背板属(Teladorsagia)为代表2-2-1模式(见图2A),另一类以奥斯特属(Ostertegia)为代表2-1-2模式(见图2B),马歇尔属(Marshallagia)交合伞宽大,伞肋较长,但侧肋模式与奥斯特属相同,图3A显示马歇尔属线虫交合伞及其肋和乳突数字系统[7]。奥斯特亚科中,生殖锥详细结构不易观察,但仍可根据其生殖锥结构共有特征鉴别[7]:其一,腹面带有两个细小乳突(“0”乳突)(见图3A);其二,背面带有两个被副伞膜围绕、较长而纤细乳突(“7”乳突,也称背小肋)(见图3A、B、C)。生殖锥结构代表奥斯特亚科属一级水平重要差别[3],奥斯特属生殖锥腹面前缘有明显突起(见图3B),背板属该部位无突起(见图3C、D)。奥斯特亚科种多态性研究为种形态学鉴别提供新思路,在多态性种中,虽然雄虫形态不同,但其主级与次级形态类型间差别模式一致[6-7],所有主级形态类型均具细长交合刺,生殖锥背面存在未角质化副伞膜(见图3B、C),而次级形态类型交合刺相对短粗,生殖锥背面显示一个扩大且呈骨质化倾向板状结构(见图3D)。生殖锥形态差别最典型例子是环纹背板线虫(T.circumcincta)(主级形态类型)(见图3C,图4A)、“三角背板线虫”(T.trifurcata)(见图3D)(次级形态类型)、“达氏背板线虫”(T.davtiani)(次级形态类型)(见图4B),均属于同一虫种三种不同形态类型。因此,仅凭交合刺长度及单一雄虫形态差别鉴定虫种标准难以普遍适用于毛圆科线虫形态分类。

图2 奥斯特亚科交合伞侧肋模式Fig.2 Pattern of lateral raysof Ostertagiinae
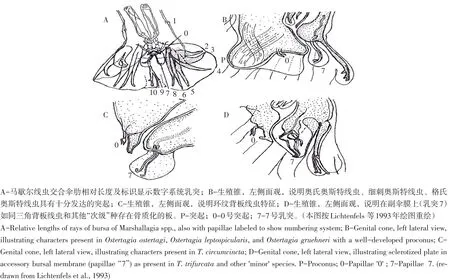
图3 奥斯特亚科交合伞和生殖锥特征Fig.3 Drawingsillustrating characteristicsof copulatory bursae and genital cones of Ostertagiinae

图4 背板线虫生殖锥(背、腹面观)(按Skriuabin等1954年绘图重绘)Fig.4 Genital cone of Teladorsagia spp.(dorsal and ventral view)(redrawn from Skriuabin et al.,1954)
2 奥斯特亚科属和种系统分类学
2.1 传统系统分类学
Skriuabin等经典分类学,将奥斯特亚科线虫归于毛圆亚科(Trichostrongylinae)奥斯特族(Ostertagiea),共15属3亚属[3]。Gibbons等认为奥斯特亚科有17个属[10],但Durette-Desset等认为有5或6个属[9,11],Jansen认为有7个属[12]。自Dróżd ż提出多态性假说以来[13],分类观点一直存在争议,亚科系统分类学及种间关系复杂。Lichtenfels等建议根据共有可鉴别特征或杂交试验建立分类单元属[5];奥斯特亚科虫体生殖锥腹面成对“0”乳突代表1个共有可鉴别特征,一些缺乏特征属如Gazellostrongylus可从该亚科移出。Lichtenfels等根据北美野生和家养反刍动物寄生线虫种类、地理分布及形态学特征,列出奥斯特亚科种检索表,将雄虫生殖锥结构、伞肋模式作为主要检索特征,雌虫在多数情况下根据体表纵嵴模式和体中部嵴数目、食道与小肠间阀门长度鉴定至种;建议奥斯特亚科包括8属[5]:奥斯特属、马歇尔属、蛇圆属(Camelostrongylus)、长圆属(Longistrongylus)、Talandorsagia、刺翼属(Spiculopteragia)、短角鹿圆属(Mazamastrongylus)和猪圆属(Hyostrongylus)。
Dróżd ż认为次级变体发生或缺乏属一级特征,每个多态性有一个特定类型次级变体[4]。根据有无多态性将奥斯特亚科内属归纳为9个属:奥斯特属/斯纳属(Skrijabingia)(次级变体)、马歇尔属/粗交合刺属(Grosspiculagia)、奥洛夫属(Orloffia)/Buriatica(次级变体)、背板属/Trifurcata(次级变体)、刺翼属/里纳德属(Rinadia)/Apteragia(次级变体)、蛇圆属、短角鹿圆属、Sarwaria、长圆属缺乏次级变体。综上可知,Skrjabingia为Ostertegia同物异名,Grosspiculagia为Marshallagia同物异名,Rinadia和Apteragia为Spiculopteragia同物异名。
奥洛夫属其种此前归于奥斯特属[3,6,14]或马歇尔属[9],但Dróżd ż认为奥洛夫属种虽与奥斯特属近似,但主级变体生殖锥腹面缺乏突起,且主级变体或次级变体伞肋“5、6、8”末端等距离靠近,而奥斯特属具2-1-2伞肋结构和生殖锥腹面前缘有明显突起(次级变体突起不明显),应为姊妹属[4]。
关于奥斯特亚科属鉴定,马歇尔属交合伞宽大,后面肋比腹肋长,伞肋模式是2-1-2,易区别;蛇圆属和长圆属伞肋模式是2-1-2,前者有细长呈海绵状交合刺,后者有粗短背肋,两者可区分[9];背板属伞肋模式是2-2-1,生殖锥腹面前缘无突起,与奥斯特属区别前文已述;短角鹿圆属和刺翼属2-2-1伞肋模式,两属通过交合刺、副伞膜和体表纵嵴结构可区分[3,15]。Lichtenfels和Dróżd ż等认为短角鹿圆属和刺翼属为姊妹属,两个属仅来自鹿科动物且完全平行进化,前者为单形态性而后者有“Rinadia”样次级变体发生[7,16]。Sarwaria属仅包含一个种,即S.bubalis;Jansen将其归于短角鹿圆属,但交合伞、副伞膜和交合刺结构与该属不同[15];该虫种是亚洲水牛(Bubalusbubalis)和瘤牛(Bosindicus)寄生虫,而短角鹿圆属虫种来自北美鹿科动物寄生虫,故Dróżd ż仍将S.bubalis保留在Sarwaria属[4]。
2.2 分子生物学分类
随着分子生物学发展,以DNA为基础分子标记技术备受关注,包括限制性片段长度差异(RFLDs)、扩增片段长度多态性(AFLP)、限制性片段长度多态性(RFLP)、DNA随机扩增多态性(RAPDs)、核糖体(rDNA)内部转录间隔区(ITS)、单序列重复/微卫星(SSR)及线粒体DNA(mtDNA)cox1或Nad4位点分析等[17-18]。Blouin对包括背板属、奥斯特属在内7个圆线虫属20种虫体样品作ITS和mtDNA Cox1和Nad4基因分析,发现mtDNA基因比ITS区具有更快突变率,更适于少量个体中确定隐藏种[18]。对于奥斯特亚科虫种,rDNA ITS-1和ITS-2序列分析已用于种验证、新种和错定种识别。Stenvenson等首次使用rDNA ITS-2标记,分析环纹背板线虫、三叉背板线虫、达氏背板线虫、奥氏奥斯特线虫(O.ostertagi)和细刺奥斯特线虫(O.leptospicularis)样品,证明文献记载背板属3个种在该序列上无显著差异,背板属和奥斯特属间有13%~15%序列差异[19]。格氏奥斯特线虫(O.gruehneri)和来自驯鹿O.arctica的ITS-1和ITS-2序列分析表明,地理分布相隔遥远的奥斯特线虫存在种多态性[20]。Dallase等对来自挪威和欧洲冻土地带驯鹿线虫样品的rDNA ITS-1和ITS-2序列分析表明,所检线虫样品在该序列上几乎相同,结果证明O.gruehneri和O.arctica为同一物种二态型雄虫[20]。Dallas等用上述相同分子标记,证明马氏马歇尔线虫(M.marshalli)和西方马歇尔线虫(M.occidentalis)为同一物种二态型雄虫[21]。Santin-Duran等为证实刺翼属多态性种假说,以rDNA ITS-1和ITS-2及mtDNA中Nad4为分子标记,证明Spiculopteragia mathevossiani为S.spiculoptera次级形态变体[22]。Lienard等用相同分子标记证明,S.quadrispiculata为S.asymmetrica次级形态变体[23]。Sultan等研究亚洲鹿S.houdemeri和亚洲野牛S.andreevae样品,经形态学鉴定,使用ITS和部分mtDNA Co1为标记,发现雄虫和雌虫均有相同遗传学背景,证明S.Houdemeri存在多态性,即S.andreevae为S.houdemeri次级形态变体[24]。Grillo等使用多位点群体遗传学技术,分析环纹背板线虫、三叉背板线虫及达氏背板线虫各31种个体、5个微卫星基因型,证明3种线虫间无遗传学差异,支持三叉背板线虫、达氏背板线虫为环纹背板线虫次级形态类型假说[25]。Hoberg等在北美麝牛体内发现曾被误认为环纹背板线虫的一个隐藏种,经mtDNA Nad4基因序列测定证明隐藏种与环纹背板线虫线粒体DNA Nad4基因序列差异为13%[26],命名为北极背板线虫(T.boreoarticus)。Lv等对来自中国北方地区奥斯特亚科3个属(背板属、马歇尔属、奥斯特属)11个虫种,首次用DNA条形码技术分析mtDNA Co1基因,表明Co1基因在种间双参数遗传距离(K2P)(13.6%)是种内差异(3.69%)近4倍[27]。Neighbor-Joining法系统进化分析显示12个分支,且环纹背板线虫序列分2支,2支平均K2P达12.1%,表明该线虫可能是种复合体;该结果与微卫星基因型结论不同,需更丰富分子生物学数据研究环纹背板线虫群体遗传学特征。
3 我国奥斯特亚科分类学研究现状
3.1 国内发现奥斯特亚科线虫新种及分类地位
自50年代初至80年代末,国内学者在各地开展家养动物寄生虫区系调查和分类鉴定并取得重要成果。目前,奥斯特亚科中,国内共报道21个新种,大多来自家养反刍动物,仅3个来自羚牛、岩羊和藏原羚,表1列出我国20个新种发现地点、宿主等情况(短肋奥斯特线虫除外)。
从表1可知,熊氏奥斯特线虫和叶氏奥斯特线虫均来自甘肃武威县羊体内,2种线虫属于奥斯特属,从许授泰等发表新种时绘图和描述可知[28],生殖锥“7”乳突周围呈弱角质化或板状,根据次级变体形态学鉴定依据,两个种可能是奥斯特属某个主级种次级形态类型,或一个是主级种,另一是次级变体。关于异刺奥斯特线虫,原描述仅以一条雄虫为代表,未见雌虫,由许授泰等绘图可见[29],该种虫体两个交合刺长度差异较大,与属特征不对应,可能为奥斯特属其他种次级变体或畸形,无法作为独立种。沈杰认为吴兴奥斯特线虫与长刺奥斯特线虫为同一种[46],鉴于前者发表年代较早,故为有效种。中华奥斯特线虫和西藏奥斯特线虫标本来源于同一地区且宿主相同,对比孔繁瑶等发表新种[33-34],本文认为形态无差异,应为同一种;另外,原文记载雄虫数量少,且与其他2个未知种同时寄生,雌虫无法鉴别,绘图中2个种交合伞肋模式为2-2-1结构,交合刺细长,应归于背板属非奥斯特属;根据发表时间,后者应为前者次级同物异名,但有效性有待考证(参见文内3.2)。个别文献记载短肋奥斯特线虫(O.shartdorsalray),伞肋式与背板属相同应归背板属非奥斯特属,仅背肋短无法构成新种条件,且环纹背板线虫具多态性,雄虫形态差别较大,故该种应为环纹背板线虫。念青唐古拉奥斯特线虫定种时位于粗交合刺亚属,由孔繁瑶等发表新种原文描述和绘图可知,从藏羊皱胃中发现1 000多条奥斯特线虫,且与奥氏奥斯特线虫(O.ostertegi)联合寄生,数量仅9条,雄虫交合刺粗短,根据多态性假说,该种可能为奥氏奥斯特线虫次级形态类型[32]。宁陕奥斯特线虫命名时位于粗交合刺亚属,从绘图上看该种交合伞符合2-2-1模式,应属背板属,且交合刺粗短,可能为背板属中某一个种次级形态[36]。矛形奥斯特线虫定种时位于粗交合刺亚属[37],因粗交合刺亚属是马歇尔属同物异名或次级变体[4],符合马歇尔属特征,定种时未与马歇尔属其他种比较,该种与青海马歇尔线虫[43]均来源于青海祁连县,宿主均为岩羊,前者应属马歇尔线虫属,由于定种时两种位于不同属,后者定种时未与前者比较,从雄虫交合伞、交合刺特征看,两者差异不明显,应为马歇尔属同一种。甘肃奥斯特线虫和粗刺马歇尔线虫,前者由陈义民定种[35],位于粗交合刺亚属,交合刺粗大,从绘图看伞肋式为2-1-2符合马歇尔属,因粗交合刺亚属为马歇尔属同物异名,故该种隶属马歇尔属非奥斯特属;后者由李丛林等定种[42],定种时两种未归同属,且后者未与前者比较,两者特征如交合刺结构和长度接近,应属同一个种;此外,粗刺马歇尔线虫为蒙古马歇尔线虫(M.mongolica)次级形态类型[47],甘肃奥斯特线虫亦如此。塔里木马歇尔线虫[40]、天山马歇尔线虫[39]及短尾马歇尔线虫[41]均来自新疆,宿主为绵羊,交合刺差异较小,可能为同一个种。拉萨马歇尔线虫[44],与蒙古马歇尔线虫尺寸相近,引带形状不同,但其他形态与蒙古马歇尔线虫无本质区别;由于本属线虫雄虫多态性,例如西方奥斯特线虫(O.occidentalis)和马氏马歇尔线虫(M.marshalli)是同种不同形态类型[47],故本文初步认为拉萨马歇尔线虫为蒙古马歇尔线虫同物异名。斯纳属为奥斯特属同物异名,但四川斯纳线虫,伞肋式符合2-2-1排列,应属背板属[45],背板属线虫在野牛中报道仅有北极背板线虫[48],四川斯纳线虫寄生于黄牛皱胃,可能为背板属中一个隐藏种或环纹背板线虫次级形态。对于斯氏奥斯特线虫[30]、新疆马歇尔线虫[38]及祁连马歇尔线虫[43],本文保留原有分类和定种依据。以上评述未见原始描述标本,仅以形态判定,尚待考证。
3.2 当前研究现状

表1 国内报道奥斯特亚科新种Table 1 New species of Ostertagiinae reported in China

续表
我国反刍动物寄生奥斯特亚科线虫有3个属,分别是马歇尔属、背板属、奥斯特属,广泛发生且明确存在。但目前有效种尚不清楚,其他属是否存在有待深入研究。
目前,国内寄生虫系统分类学资料报道很少,尤其是毛圆科奥斯特亚科种多态性、隐藏种等。2004年宁夏银川地区绵羊真胃中曾发现环纹背板线虫(曾称为环纹奥斯特线虫)和三叉背板线虫联合寄生情况(未发表资料),其雄虫交合刺如图5(A、B)所示,其中1只绵羊真胃中收集约600条背板线虫,包括环纹背板线虫雄虫150条,三叉背板线虫雄虫17条,前者占89.8%,后者占10.2%,是典型多态性种联合发生案例。2009年,在兰州肉联厂获得的绵羊真胃中发现少量疑似环纹背板线虫寄生,但雄虫交合刺细长,其尖端部分与中华奥斯特线虫或西藏奥斯特线虫相似(见图5C),同时也有少量三叉背板线虫雄虫(未发表资料),说明我国存在该属线虫多态性现象,但主级种雄虫交合刺形态差异国外未见报道,目前标本数量极少,无法确定。
西北民族大学生命科学与工程学院动物医学教研室在宁夏地区收集绵羊真胃标本中,获得少量蒙古马歇尔线虫,同时发现比蒙古马歇尔线虫交合刺粗短的1条雄虫(见图6A、B)(未发表资料),因标本数量极少,无法确定。现认为粗交合刺马歇尔线虫为蒙古马歇尔线虫次级形态类型,上述雄虫虫体尺寸等形态特征与蒙古马歇尔线虫无明显差异,仅交合刺为蒙古马歇尔线虫雄虫2/3,可能为蒙古马歇尔线虫另一种次级形态类型。
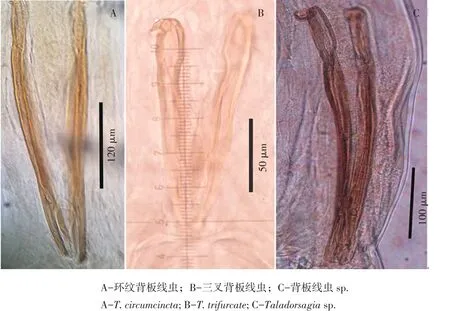
图5 背板线虫雄虫后端Fig.5 Posterior ends of Teladorsagia spp.
以上发现表明我国存在奥斯特亚科线虫种多态性,但对毛圆科线虫,特别是奥斯特亚科系统分类学进展及种多态性、隐藏种及杂交等研究较少。国内仍沿用50年代分类系统,分类标准混乱[34,49-50],例如环纹背板线虫和三叉背板线虫,已隶属背板属,国内至今仍沿用50年代旧名普通奥斯特线虫或环纹奥斯特线虫和三叉奥斯特线虫。1954年因发现“达氏背板线虫”建立背板属,这3种线虫为同一种不同形态类型,即根据多态性假说,后2个种为环纹背板线虫2种次级变体。
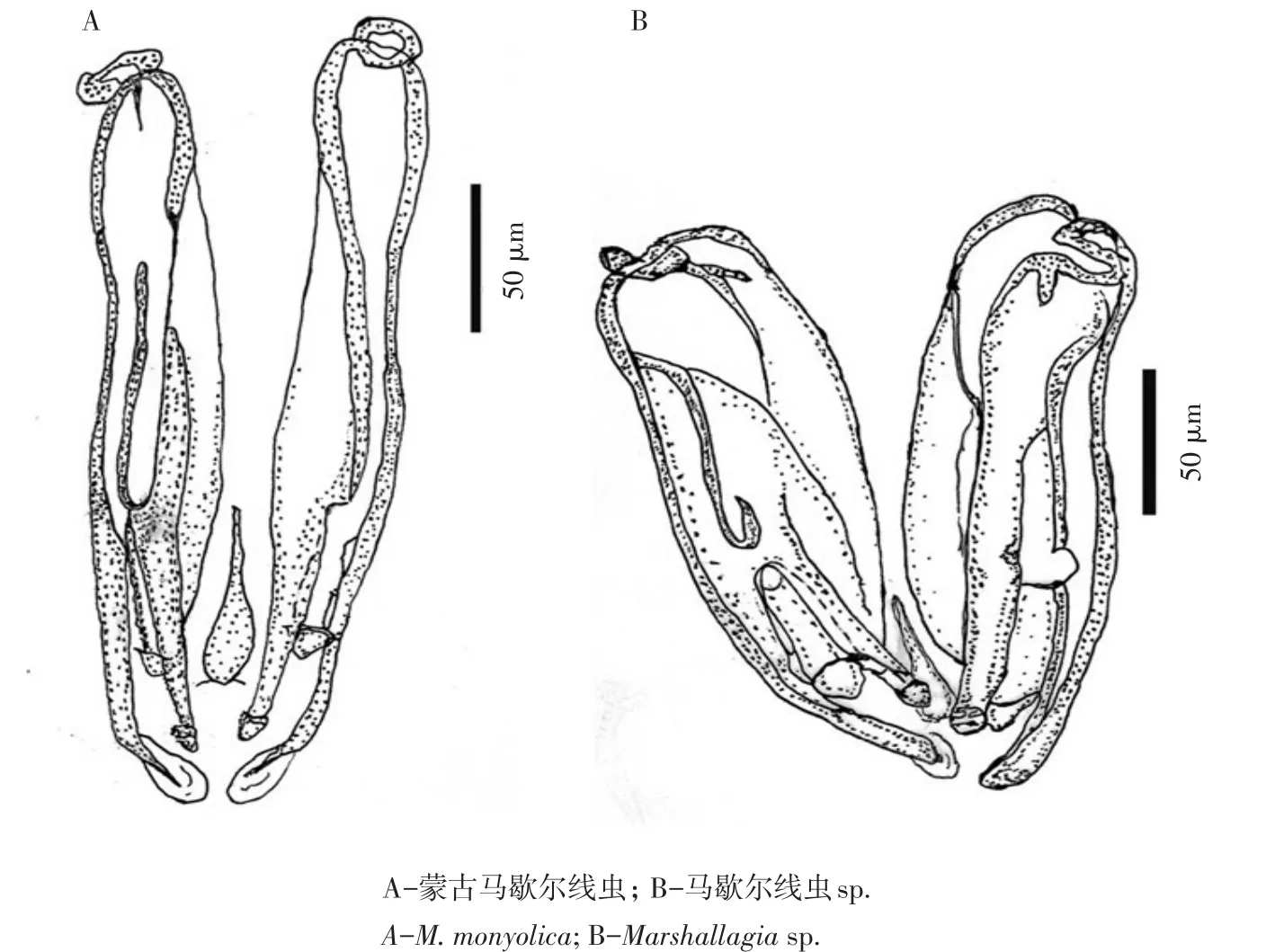
图6 马歇尔线虫交合刺绘图Fig.6 Drawingsof spiculesof Marshallagia spp.
4 结论
随着电子显微镜和分子生物学技术在生物科学包括寄生虫分类学中的应用,可解决该亚科线虫系统分类学、属和种鉴定错误、种分类地位等问题。新技术应用可阐明形态学细微结构,并将阐明结构与系统发育间关系。奥斯特亚科线虫系统分类学还涉及该亚科线虫生物学、历史地理、种多态性和隐藏种、群体遗传学及宿主与寄生虫间互作等,尚需全面研究,阐明相互关系。鉴于目前我国在毛圆科奥斯特亚科线虫系统分类学研究现状及与国外差距,建议全面开展我国反刍动物(包括家养和野生)相关线虫调查,继续收集新样品并结合以往标本,在形态分类学基础上,利用先进研究手段,对标本作详细形态学描述和遗传学研究。加强寄生虫分类学人才培养,修订我国毛圆科线虫奥斯特亚科属和种分类标准。
猜你喜欢
甘肃农业科技(2021年6期)2021-07-25
物理之友(2020年12期)2020-07-16
武夷科学(2019年2期)2019-12-20
科学导报(2019年35期)2019-09-03
西北农林科技大学学报(自然科学版)(2018年5期)2018-05-24
幸福·婚姻版(2018年3期)2018-03-22
武夷科学(2017年0期)2017-03-10
剑南文学(2016年14期)2016-08-22
军事历史(1984年2期)1984-08-21
