
凤丹灰霉病病原鉴定与发病条件研究
2018-09-11 08:32:50罗长维
河南农业科学 2018年8期
罗长维,陈 友
(1.重庆城市管理职业学院,重庆 401331; 2.中国林业科学研究院 资源昆虫研究所,云南 昆明 650224)
凤丹(PaeoniaostiiT.Hong & J.X.Zhang)隶属于芍药科(Paeoniaceae)芍药属(Paeonia)牡丹组(Sect.Moutan),落叶小灌木,为杨山牡丹变种,因产于安徽铜陵的凤凰山和南陵西山得名[1]。凤丹花冠洁白硕大,单花顶生,肉质根皮肥厚,经干燥可制成中药丹皮。芍药属植物主要分布于中国,大部分种类雄蕊瓣化严重,结籽少或不结实;少数种类为单瓣花冠,结实量较大,如凤丹。凤丹蓇葖果具5~7枚单果,每果黑色种子数可达50粒。种仁富含具“血液营养素”美称的α-亚麻酸,含量高达42%,远高于其他食用油,为我国近年来重点发展的优质食用油原料[2-3]。由于结果率高、种子量大、生态适应性广等优点,且兼具观赏、药用价值,凤丹在我国山东、河南等黄河流域及重庆、湖南等长江流域均有广泛种植[1-2]。尽管凤丹籽油品质优良、种植面积大,由于存在病害严重以及缺乏籽油良种等问题,种子产量无法满足凤丹籽油产业化发展的需求。
灰霉病是世界上最具毁灭性的植物病害之一,由半知菌亚门的葡萄孢真菌(Botrytisspp.)与类葡萄孢真菌(Botrytis-like fungi)引起。国外对灰霉病研究比较深入,在病原菌生物生态学特性、致病机制、抗病性诱导以及综合防治等方面均取得了较大进展[4]。我国对灰霉病的研究则相对滞后,以化学防治的药效研究为主,也有关于其病原菌生物学特性、抗药性等方面的报道[4-7]。灰霉病也是我国凤丹生长期的主要病害,严重影响其结果率与种子产量。目前,有关凤丹的研究主要集中于良种选育、籽油化学组成和加工等方面,尚无有关凤丹灰霉病的报道[3]。对芍药属其他植物灰霉病的报道表明,牡丹灰霉病在我国的山东、河南等地,以及牡丹分布的欧洲西部与美洲均普遍发生[5-11],主要由灰葡萄孢(B.cinereaPers,有性型BotryotiniafuckelianaWhetzel)、牡丹葡萄孢(B.paeoniaeOudem.)或假灰葡萄孢(B.pseudocinere)侵染引起[8-11]。其中,灰葡萄孢为一种泛化性的兼性寄生真菌,出现在200多种植物上[4]。该病原菌主要靠分生孢子随气流、雨水以及农具等传播扩散[7],防治较为困难。牡丹葡萄孢目前仅在芍药属植物上发现,危害程度相对较轻[11]。假灰葡萄孢常常与葡萄孢真菌伴生,在葡萄、牡丹等多种寄主植物的灰霉病病原菌中均有发现,危害通常较小[10-11]。基于此,本研究对凤丹灰霉病病原进行鉴定,并对其田间发病规律与流行特点进行分析,以期了解该病的发生危害条件,为下一步病害的有效防控提供实践依据和理论参考。
1 材料和方法
1.1 试验地概况
试验点位于重庆市垫江县长龙镇(107.406 42°E、30.305 57°N)。垫江县地处华蓥山脉东部,地貌以丘陵为主,海拔320 ~1 183 m,年均温16 ℃,年均日照 1 260 h,年均降水量1 160 mm,为中亚热带季风性湿润气候。垫江为我国中药材丹皮的主产区之一,也是重庆凤丹主要种植地,灰霉病危害较重。试验点凤丹居群株龄约为5 a,株高1.0~1.2 m,株行距(0.6~0.8)m×(0.8~1.0)m。
1.2 研究方法
1.2.1 病原菌鉴定 于2017年4月采集凤丹灰霉病感病器官(花果、茎叶)30份,分株分类装入纸袋,在实验室内通过组织分离法对病原菌进行分离与纯化。将供试菌株移植于PDA培养基平板上,置于20 ℃恒温箱培养,在显微镜下对病原菌的培养性状及分生孢子梗、分生孢子、菌核等形态特征进行观察。
1.2.2 病原菌越冬分析 于2016年7月采集凤丹感病部位(花、果、茎、叶),干燥后置于硫酸纸袋内,分别存放于室内、置于室外干燥通风处、置于室外地表(纱网袋内)以及埋于土壤里(深度10 cm),于次年3月取出样本进行病菌分离培养,并在显微镜下进行观察。
1.2.3 流行规律及发病条件分析 于2015—2018年的3—6月选择有代表性地块约1 hm2作系统监测田,定点、定株、定期调查灰霉病田间流行动态与发病条件。采用平行线取样法取5个样点进行调查,每点选标准株20株,调查花果发病情况,重复3次。前期调查表明,凤丹灰霉病基本上仅危害花果,极少危害茎干、叶片,因此灰霉病发病情况的调查依据为花果是否感病。其中,病情按花朵或果实病斑面积占整个花朵或果实面积比例来分级:0级—无病斑,1级—病斑面积5%以下,3级—病斑面积6%~10%,5级—病斑面积11%~20%,7级—病斑面积21%~50%,9级—病斑面积50%以上[12]。发病率与病情指数计算公式如下:


1.3 数据分析
试验结果用平均值±标准误表示。采用SPSS 22.0软件对凤丹灰霉病的发病率与病情指数进行单因素方差分析(ANOVA)。
2 结果与分析
2.1 凤丹灰霉病的病原鉴定与危害特征
经形态鉴定,凤丹灰霉病的病原为灰葡萄孢(B.cinereaPers),属半知菌亚门(Deuteromyeotina)、丝孢纲(Hyphomycetes)、丝孢目(Hyphomycetales)、淡色孢科(Moniliaceae)、葡萄孢属(Botrytis)。其菌丝体表生,分生孢子梗直立,数根丛生,分生孢子多为椭圆形或卵形,少数球形,在分枝末端聚生成葡萄穗状,淡褐色至无色,单胞。灰葡萄孢菌为兼性寄生真菌,能以菌丝、分生孢子及菌核的形式感染,以坏死组织为寄主,能杀死植物细胞,并在植物的坏死组织上生长发育。灰葡萄孢菌群在形态上有所差异,可能来自于不同的菌群亚型。病菌危害部位初期产生透明的点状病斑,之后扩大呈不规则形,长出灰色菌丝。病部菌丝发展到一定阶段时,可产生大量分生孢子梗,其上聚生葡萄穗状分生孢子,使病部呈灰霉状,病斑处有时产生黑色颗粒状的菌核。
凤丹灰霉病主要危害幼嫩花果,包括花蕾(图1A)、花朵(图1B)、幼果(图1C)及中果(图1D),极少感染嫩叶与茎干。一般从花萼开始感病,病斑逐渐从花萼端部扩展至花萼基部,随后侵染花瓣基部与花梗,花瓣与花蕊随之感病。由于许多花朵生长略有倾斜,雨水易积部位常常先受害。由于田间湿度大,感病花瓣无法干燥并脱落完全。残留花瓣黏附在子房基部,成为幼果灰霉病侵染源。灰霉病发病迅速,花果一旦感病,感病部位(花瓣、花药以及掉落在花瓣上的花粉、幼果)很快变褐、软腐。

A.感病花蕾;B.感病花朵;C.感病幼果;D.感病中果图1 凤丹灰霉病感病症状
2.2 凤丹灰霉病的发生规律
2.2.1 病原菌的越冬 病原菌越冬检测结果表明,在室内存放和悬挂于室外的花果病残体中均未分离到灰葡萄孢;位于地表的花果病残体与埋于土表层下10 cm的花果病残体中也始终未能分离到该病原菌。病残体上仅分离到1种专性腐生真菌链格孢菌[Alternariaalternata(Fr.) Keissler],可能为灰霉病感染后期侵入的病原菌。上述观测结果表明,灰葡萄孢在凤丹果实上发病完成生活史后,无法在感病花果上继续生存,后期可能主要以菌核残留在土壤中越夏、越冬。
2.2.2 田间流行动态 凤丹灰霉病从3月上旬开始发生。3月中旬至下旬,随着气温的上升,花朵发病率呈上升趋势,但差异并不显著。花朵发病率在3月上旬与4月上旬之间差异显著。从4月上旬开始,日均温上升到19 ℃,发病率呈现快速增长,4月上旬、4月下旬与5月上旬之间发病率呈现显著差异。5月上旬后,发病率不再上升,5月上旬、5月中旬、5月下旬与6月上旬之间发病率均无显著差异(图2)。
病情指数与发病率在5月上旬前存在正相关。凤丹灰霉病致病性强,花果一旦被感染,快速引发宿主软腐,引起成熟或衰老组织的严重破坏,导致其腐烂,因此凤丹发病率与病情指数在前期密切相关。从3月上旬到4月上旬,病情指数随时间呈缓慢上升趋势,但无显著差异。病情指数在3月上旬与4月中旬之间差异显著。随着4月气温升高,病原菌增殖周期缩短,病情指数上升速度较快,4月中旬与5月上旬之间病情指数具有显著差异。5月上旬后,病情指数呈下降趋势,且5月上旬、5月中旬、5月下旬与6月上旬之间的病情指数无显著差异(图2)。
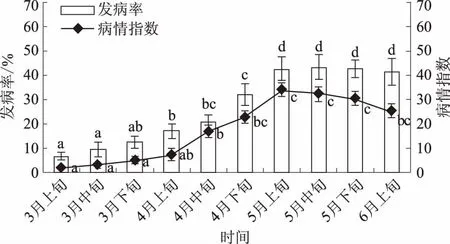
不同字母表示差异显著(P<0.05),下同图2 凤丹灰霉病发生动态
2.3 环境因子对凤丹灰霉病发病的影响
2.3.1 年份 从图3可以看出,凤丹灰霉病年年发生,但发病率与病情指数在不同年份存在显著差异。
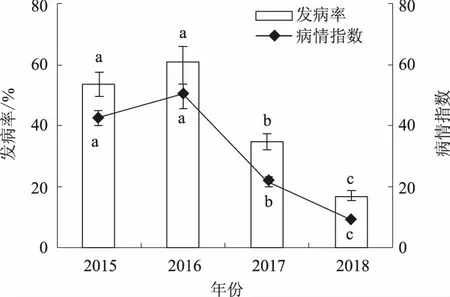
图3 年份对凤丹灰霉病发病的影响
其中,2016年凤丹感病最重,发病率与病情指数分别为61.10%、50.22。在2015年,凤丹发病率与病情指数分别为53.33%、42.35,发病情况与2016年相比差异不显著,但病情稍轻。2017年凤丹发病较轻,发病率与病情指数分别为35.00%、22.19。2018年凤丹发病最轻,发病率与病情指数分别为17.00%、9.20。
2.3.2 花朵开放时期 从图4可以看出,不同时期开放的花朵,其发病率与病情指数有显著差异。3月20日(早期)开放的花朵,发病率与病情指数分别为25.00%、12.19,感病最轻,大部分花朵能发育为成熟蓇葖果;3月25日(中期)开放的花朵,花朵发病率与病情指数分别为36.33%、22.35,发病情况与早期相比明显加重;4月1日(晚期)开放的花朵,发病率与病情指数分别为58.10%、49.22,发病最重。

图4 花朵开放时期对凤丹灰霉病发病的影响
2.3.3 大田地势 凤丹灰霉病的发生也受大田地势影响。丘间谷地地势最低,发病最重,发病率与病情指数分别为56.67%、50.92,其次为丘陵坡地,发病率与病情指数分别为27.50%、24.55,随后为丘陵阶地,发病率与病情指数分别为22.17%、18.34,丘陵顶部最低,发病率与病情指数分别为9.17%、4.11。总体上不同地势之间差异显著(图5)。
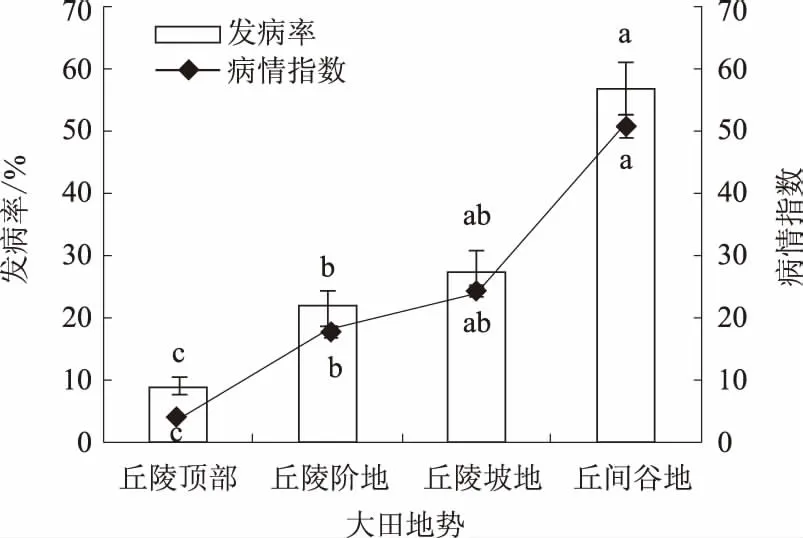
图5 大田地势对凤丹灰霉病发病的影响
2.3.4 土质类型 凤丹灰霉病的发生还受土壤质地的影响。凤丹在黏土中受害最重,发病率与病情指数分别为60.5%、49.65,壤土中受害其次,发病率与病情指数分别为31.83%、19.69,在排水良好的石砾土(石谷子地)中受害最轻,发病率与病情指数分别为19.17%、12.12。3种土质类型之间差异显著(图6)。
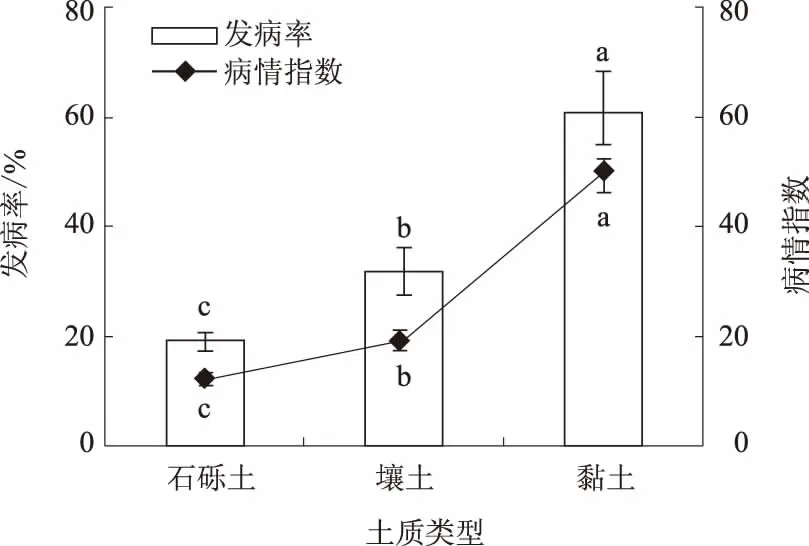
图6 土质类型对凤丹灰霉病发病的影响
3 结论与讨论
通过4 a定点试验研究,结果表明:(1)凤丹灰霉病病原菌为半知菌亚门的灰葡萄孢;(2)凤丹灰霉病为低温高湿型病害,发生规律与气候因子密切相关;(3)不同年份、花期、大田地势与土质类型对灰霉病发病都有一定的影响。
凤丹灰霉病病原菌为半知菌亚门的灰葡萄孢。对于芍药属植物灰霉病的病原菌,各地报道不一。欧美等地报道牡丹灰霉病病原主要有灰葡萄孢、牡丹葡萄孢和假灰葡萄孢3种[11,13],其中灰葡萄孢危害最重,牡丹葡萄孢危害其次,假灰葡萄孢危害最轻[11]。我国对芍药属植物灰霉病病原的研究结果差异较大。杨瑞先等[7]与陈士宁等[14]分别报道河南洛阳的牡丹与北京的芍药灰霉病病原菌为灰葡萄孢。薛杰等[5]和易图永等[15]分别报道山东与湖南两地牡丹灰霉病的病原菌为牡丹葡萄孢。杨瑞先等[6]随后通过RAPD方法对牡丹灰霉病病原菌进行分子生物学鉴定,证实其为灰葡萄孢,且菌群类型多样。本研究对重庆垫江凤丹灰霉病感染的花果与茎叶进行采集,采集时间位于灰霉病发病中期,均只发现灰葡萄孢,未发现牡丹葡萄孢,这与杨瑞先等[6-7]、陈士宁等[14]的鉴定结果一致,与薛杰等[5]、易图永等[15]的鉴定结果不同,与欧美等地病原菌差异较大。因此,凤丹灰霉病的病原是否有牡丹葡萄孢,还有待进一步观测研究。
凤丹灰霉病为低温高湿型病害,发生规律与气候因子密切相关。灰葡萄孢菌的生长适温是16~24 ℃,超过20 ℃产孢能力显著降低,超过24 ℃则菌株生长缓慢[14]。许多学者对草莓、番茄等作物的灰霉病发生规律进行研究,结果表明,灰霉病一般在阴雨潮湿的天气容易出现发病高峰[16-17]。重庆大部分地区初春低温多雨低光照,凤丹生长势弱,细胞膨压低,抗病性差,而病菌仍能萌发侵染,因此低温多雨低光照的初春有利于分生孢子的大量形成和传播,植株易受病菌感染。5月上旬后,气温升高、空气湿度降低、光照增加,对灰霉病菌的萌发与生长及产孢量均具有抑制作用,发病率虽未下降,但病情指数逐渐降低。此外,湿度的昼夜变化对灰霉病发生也有影响,早晨由于温度的逐渐升高与湿度的下降使分生孢子释放于周围空气中,导致分生孢子扩散,而夜间的高湿则有利于灰葡萄孢的萌发与生长[14]。
不同年份、不同花期对灰霉病发病都有一定的影响。年份不同,降雨量与温度存在差异,植株发病率也随之变化[18]。在2015年、2016年、2017年与2018年的2—4月,垫江县的降雨量分别为230.2 mm、260.8 mm、173.0 mm与190.6 mm。灰葡萄孢菌分生孢子随降水、空气等传播,尽管2018年降雨量与2017年差异不大,但降雨基本发生在夜间,白天常为晴天,因此发病最轻。而2016年降雨量最多,因此灰霉病流行,发病也最重。降雨量大、光照不足时也会引起田间湿度大,植株生长弱,利于病原菌的萌发与生长[18]。尤其不利的是,感病花瓣在田间湿度较大时不易脱落,常常残留在幼果基部,在增加幼果基部空气湿度的同时,也明显增加了病菌数量,从而增加幼果感病率。相反,如果田间湿度低,花瓣易干燥脱落,花果感病则较轻。此外,凤丹花朵开放时间越晚,感病则越重。一方面,开花较晚的花朵通常受限于营养供给,生长发育缓慢、抗病能力较弱;另一方面,由于清明节的到来,降水量逐渐增加及田间湿度提高,适于灰霉病病原菌的增殖与传播,因此灰霉病发生更为严重。
大田地势与土质对灰霉病的影响也较大。位于丘陵顶部的地块地下水位低,凤丹长势好,且丘陵顶部风大、田间湿度低,因此发病最轻。反之,丘间谷地地下水位高、土壤透气性差,凤丹长势差,而且丘间风小、田间湿度大,凤丹发病最重。生长势的差异决定着凤丹抗病性的高低。生长势良好的凤丹抗病性强,一方面通过增强机械阻挡来完成,如增强细胞膨压,相应的抗病原菌侵染能力强,另一方面则是通过增强与抗病相关的生物酶活性来提高抗病性,因而长势好的凤丹不易受害或受害轻,且受害后更容易恢复。以往很多研究也表明,灰霉病、灰斑病等多种植物病害皆易发生于洼地或谷地等地势低矮之处[16,18]。在3种土壤质地中,黏土种植的凤丹发病最重,石砾土种植的凤丹发病最轻。种植在石砾土里的凤丹根系正常,而种植在黏土里的凤丹其根颈处的表皮普遍存在软腐。这表明由于黏土含空气较少,凤丹根系呼吸作用受到抑制,并进一步影响其养分吸收与植株生长势。可能与凤丹根系为肉质根,耐干旱而不耐水湿,喜干燥疏松、排水良好的土壤有关。
致谢: 感谢洛阳理工学院杨瑞先副教授与西南大学周彤副教授在凤丹灰霉病病原菌鉴定中给予的支持与帮助。
猜你喜欢
特产研究(2022年6期)2023-01-17 05:06:02
今日农业(2021年21期)2022-01-12 06:31:36
浙江林业科技(2021年3期)2021-07-13 09:03:56
植物资源与环境学报(2020年4期)2020-08-10 05:23:14
长江大学学报(自科版)(2018年14期)2018-07-17 08:37:02
天然产物研究与开发(2018年3期)2018-05-07 06:38:52
天然产物研究与开发(2018年3期)2018-05-07 06:38:43
新农业(2016年15期)2016-08-16 03:40:40
农家科技中旬版(2016年12期)2016-04-16 03:41:23
现代农业(2016年5期)2016-02-28 18:42:37
