
南方淮山炭疽病菌的致病力分化
2018-09-10 23:33:39丛子文钟步飞黄东益谢俊吴文嫱许云夏薇张荣萍黄小龙
南方农业学报 2018年3期
关键词:淮山
丛子文 钟步飞 黄东益 谢俊 吴文嫱 许云 夏薇 张荣萍 黄小龙

摘要:[目的]明确我国南方淮山炭疽病病原菌种类及其致病力分化状况,为淮山炭疽病的综合防治和抗病育种提供理论依据。[方法]采用形态学观察,结合rDNA-ITS序列比对和系统发育进化分析,对从我国南方6省(区)四川、贵州、云南、广西、福建和海南淮山种植区典型炭疽病病叶分离获得的45株炭疽病病原菌进行鉴定,并通过接种在4种不同淮山种质(D06-3、D06、D0108和D021)叶片上进行致病力检测,分析不同来源菌株的致病力差异。[结果]45株炭疽菌均鉴定为胶孢炭疽菌(Colletotrichum gloeosporioides)。45株炭疽菌对4种淮山种质材料均能致病,根据综合致病力可将45株炭疽菌分为强、中、弱3种致病类型。其中,6株强致病性菌株均来自广西,7株弱致病性菌株主要来自云南,32株中等致病力菌株主要来自贵州、福建和海南。[结论]来源于我国南方6省(区)的45株淮山炭疽病病原菌均为胶孢炭疽菌(C. gloeosporioides),其致病力分化明显,且与地理来源具有相关性。
关键词:淮山;胶孢炭疽菌;致病力;rDNA-ITS序列分析
中图分类号:S435.39;S432.44 文献标志码:A 文章编号:2095-1191(2018)03-0501-07
0引言
[研究意义]淮山(Dfoscorea opposita Thunb.)又称山药,是薯蓣科薯蓣属一年生缠绕性藤本植物,在我国栽培历史悠久,分布广泛,是一种药食兼用的高效经济作物(黄东益和黄小龙,2013;覃维治等,2014;汤洁等,2016),在历代本草及地方志中多有记载,始载于《神农本草经》,将其列为上品。炭疽病是对淮山生长危害最严重的病害之一,不仅感染淮山茎、叶和花,还能使零余子和薯块致病,严重影响淮山薯的产量和品质,制约了淮山产业的发展(熊军等,2016;郑雅超等,2016)。因此,明确炭疽病菌的种类及其致病力分化情况,并据以采取针对性的防治措施,对保障淮山产业的健康发展具有重要意义。[前人研究进展]国内外有关淮山炭疽病的研究主要集中在病原菌的分离、鉴定及防治药剂筛选等方面。朱桂宁等(2007)采用形态学观察,结合rDNA-ITS序列分析,将从广西5个淮山病区分离获得的炭疽病菌鉴定为胶孢炭疽菌(Colletotrichum gloeosporioi—des)。陈吉良(2011)通过rDNA-ITS序列比对和系统发育分析,发现海南不同淮山种植区发生的淮山炭疽病病原菌均为胶孢炭疽菌,同时评价了11种常见化学农药对胶孢炭疽菌的抑菌效果,发现7种农药对炭疽病病原菌具有一定的抑菌能力,其中多菌灵的抑菌效果最佳。陈富英(2013)评价了4种杀菌剂田问防治淮山炭疽病的药效,结果表明,10%苯醚甲环唑的防治效果最佳。覃维治等(2015)研究了6种药剂对广西桂南地区淮山药炭疽病的田间防效,发现32.5%阿米妙收杀菌剂对淮山药炭疽病的防治效果最佳。[本研究切入点]前人对淮山炭疽病病原菌鉴定的采样点仅局限于同一个省(区),而我国南方不同省(区)淮山的主栽品种不同,栽培条件各异,淮山炭疽病菌的种类和致病力是否存在差异目前仍不得而知,相关研究尚无报道。[拟解决的关键问题]对来源于我国四川、云南、贵州、广西、福建和海南南方6省(区)的淮山炭疽病病原菌进行分离、纯化,采用形态学观察,结合rDNA-ITS序列比对和系统发育进化分析对不同来源地炭疽病菌进行鉴定,通过接种于不同淮山种质检测各炭疽病菌的致病性,以明确不同省(区)淮山炭疽病菌的种类、遗传多样性及致病力分化特征,为淮山抗病种质筛选、抗病育种和病害防治提供理论依据。
1材料与方法
1.1试验材料
供试菌株:热带作物新品种选育教育部工程研究中心前期分别从四川、贵州、云南、广西、福建和海南等地采集淮山炭疽病植株病样,经分离培养、纯化获得45株菌株。供试淮山品系:D06—3、D06、D0108和D02 1,来源于海南大学薯蓣种质资源圃。
1.2试验方法
1.2.1供试菌株形态观察和致病性测定 将45株供试菌株接种至马铃薯葡萄糖培养基(PDA)上,28℃培养7-10 d,期间进行菌株日生长速度测定,观察病原菌菌落形态特征,记录菌落培养性状。
在供试4份淮山种质的生长旺盛期,采摘相同部位、大小一致的健康叶片,无菌水冲洗后,用75%酒精浸泡2 min,无菌水冲洗3~4次,用无菌吸水紙移去多余水分,再用打孔器挑取直径6 mm的菌饼接种于健康叶片上,28℃恒温保湿培养,对照用无菌PDA培养基接种,3次重复。接种7 d后检查病情,统计感病情况。
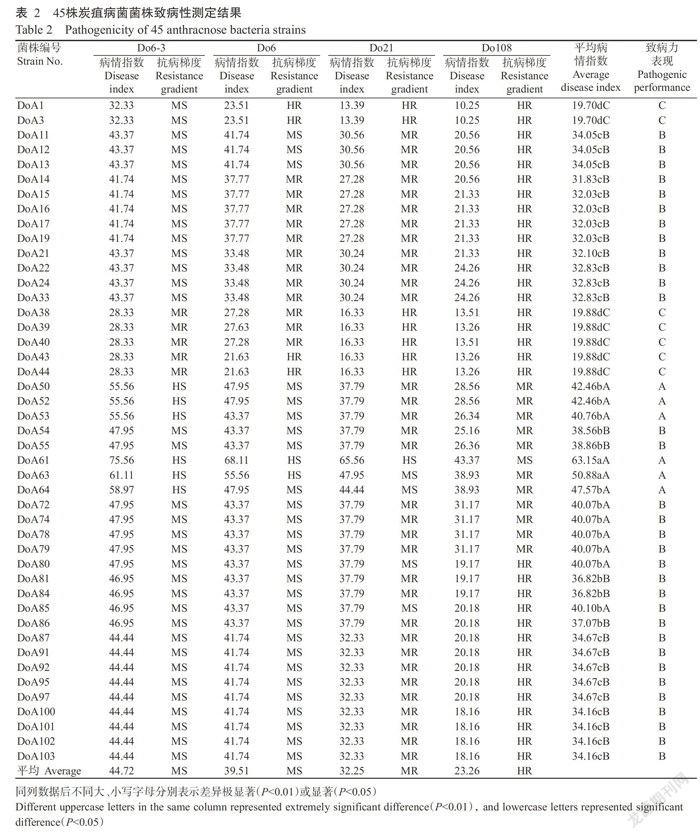
1.2.2淮山炭疽病病情分级及致病力分化测定 淮山炭疽病病情分级参照黄东益和黄小龙(2013)的分级标准:0级,接种叶片无侵染点,无病斑;1级,接种叶片病斑面积之和<1/8叶片总面积;2级,1/8叶片总面积≤接种叶片病斑面积之和<1/4叶片总面积;3级,1/4叶片总面积≤接种叶片病斑面积之和<1/2叶片总面积;4级,接种叶片病斑面积之和≥1/2叶片总面积。淮山种质对炭疽病抗病性分为5级,分级标准(王智,2004):1级,高感(HS),病情指数≥55.00;2级,中感(MS),40.00~病情指数<55.00;3级,中抗(MR),25.00≤病情指数<40.00;4级,高抗(HR),15.00≤病情指数<25.00;5级,免疫(I),0≤病情指数<15.00。菌株致病性类型划分标准(李越等,2008):强致病类型(A),平均病情指数40.00~100.00;中致病类型(B),平均病情指数20.00-40.10;弱致病类型(c),平均病指数0.10-20.10;无致病类型(D):平均病情指数为0。
病情指数=∑发病级别×相应发病级别叶数×100/(5×调查总叶数)
1.2.3淮山炭疽病菌rDNA-ITS序列分析 参考易润华等(2003)的方法,采用CTAB法提取病原菌总DNA进行rDNA-ITS序列的PCR扩增与序列分析,扩增所用引物为ITS序列通用引物ITSl(5-TCCGTAGGTGAACCTGCGG-3)和ITS4(5-TCCTCCGCTTATTGATATGC-3)。PCR反应体系25.0 LLL:10xPCR缓冲液2.5μL、2.5 mmol/LdNTP 2.0μL、2.5 u/μL TaqDNA聚合酶0.5μL、10μmol/LITS1和ITS4引物各1.0μL、DNA模板2.0μL,ddH20补足至25.0μL。扩增程序:95℃预变性3 mill;94℃1 min,55℃1min,72℃1.5 min,进行35个循环;72℃延伸10min。扩增完成后,将45株菌株PCR产物送至北京华大基因公司测序,将所得序列在NCBI数据库内进行BLAST比对分析,运用Bioedit和MEGA 4.0构建系统发育进化树。
1.3统计分析
运用SPSS 19.0进行数据统计分析,Duncans新复极差法进行差异显著性检验。
2结果与分析
2.1淮山炭疽病菌的形态学特征
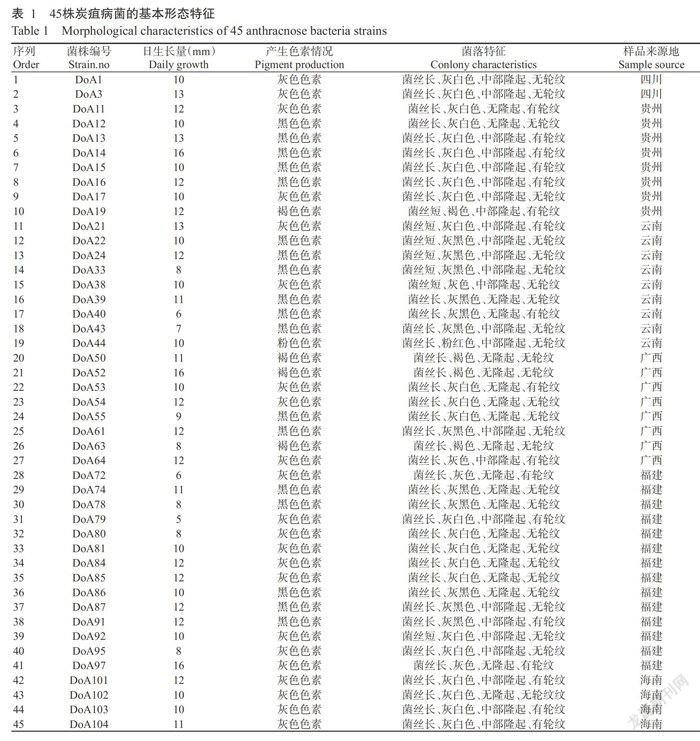
45株炭疽病菌在PDA培养基上培养5~7 d后,不同来源的病原菌培养特征存在差异(表1)。45株菌株的菌落日生长量为5~16 mm;主要产生灰色色素和黑色色素,占比分别为49%和40%,褐色色素和粉色色素较少,占比分别为9%和2%;菌丝有长和短两种形态;菌落边缘灰白色、灰黑色、灰色或褐色;菌落中部隆起或无;菌落有或无轮纹。根据病原菌上述形态特征,对照陆家云(2001)、韩长志(2012)的描述,发现45株菌株与胶孢炭疽菌(C. gloeosporioi-des)的培养特征相似。
2.2淮山炭疽病菌的rDNA-ITS序列分析结果
PCR扩增获得的45株炭疽病菌rDNA-ITS序列片段均在500~750 bp(图1),与预期结果相符。测序结果在NCBI数据库内进行BLAST比对分析,结果显示,供试的45株炭疽病菌菌株均与胶孢炭疽菌(C.gloeosporioides)具有较高的相似性,同源性在96%~100%,片段大小为520~590 bp。
从构建的炭疽病菌菌株系统发育进化树(图2)可知,45株菌株分别与已报道的胶孢炭疽菌聚在一起,但彼此的亲缘关系存在明显差异。总体上45株炭疽菌可分为两大分支,即来自福建的菌株DoA84单独构成一个分支,其他44株菌构成另一个大的分支,其中,来自海南的4株菌DoAl00、DoAl01、DoAl02和DoAl03聚在同一分支上,而其他省(区)来源的菌株相对分散地聚在不同的分支上,说明海南来源的炭疽病菌株在系统进化上具有一定的地域性,其他省(区)的炭疽病菌株地域性不强,相互问存在一定交叉。
2.3淮山炭疽病菌的致病力分化情况
45株炭疽菌对4种淮山种质材料D06-3、D06、D0108和D021均能致病,不存在专化性现象(表2);各菌株对4种不同淮山种质材料的致病力不同,有致病力分化现象。45株菌株对淮山种质材料D06-3、D06、D021和D0108的致病力依次减弱,其中,D06-3和D06表现为中感(Ms),平均病情指数分别为44.72和39.51;D021表现为中抗(MR),平均病情指数为32.25;D0108表现为高抗(HR),平均病情指数为23.26。45株菌株根据致病力分化可归为3类,即强致病性菌、中致病性菌和弱致病性菌,其中,强致病性菌共6株,平均病情指数在40.76~63.15,分别为DoA50、DoA52、DoA53、DoA61、DoA63和DoA64,均来自广西;中致病性菌共32株,平均病情指数在31.83-40.10,其中4株来自云南、8株来自贵州、14株来自福建、2株来自广西、4株来自海南;弱致病力菌7株,平均病情指数在19.70-19.88,包括来自四川的DoAl、DoA3和来自云南的DoA38、DoA39、DoA40、DoA43、DoA44。
45株炭疽病菌菌株的致病力差异变化存在一定规律性(表3)。来自四川的2株菌株均为弱致病性菌;来自贵州的8株菌株均为中致病性菌;来自云南的9株菌株中,5株为弱致病性菌,4株为中致病性菌,分别占该地区分离菌株数的55.56%和44.44%;来自广西的8株菌株中,6株为强致病性菌,2株为中致病性菌,分别占75.00%和25.00%;来自福建和海南的18株菌株均为中致病性菌。上述差异显示,在南方6省(区)淮山炭疽菌的强致病性菌主要来自广西,弱致病性菌主要来自云南和四川,中致病性菌主要来自贵州、福建和海南,菌株致病力分化与地理来源有一定关系。
3讨论
本研究采用形态学和rDNA-ITS序列分析对分离自我国南方6省(区)的淮山炭疽病病原菌进行鉴定,结果表明分离获得的45株淮山炭疽病菌均为胶孢炭疽菌(C. gloeosporioide),与朱桂宁等(2007)、陈吉良(2011)的报道一致,说明胶孢炭疽菌是我国南方淮山炭疽病的主要病原菌,该病菌在寄主上无特异性,在地理上亦无专化性。45株胶孢炭疽菌菌株问在菌落颜色、菌落形态及日平均生长量等方面呈现出多样性变化,与陈希芹(2003)有关胶孢炭疽菌菌株問形态特征上的观察结果相似。rDNA-ITS序列系统发育分析证实45株胶孢炭疽菌种内在亲缘进化关系上存在一定的遗传变异,但这种遗传变异性与菌株的形态差异及地理来源均无明显的相关性。
目前,许多研究证实植物病原菌与寄主在共同进化的过程中其致病力也随之不断变化,同时出现致病力分化现象,如大麦条纹病菌(司二静等,2015)、芝麻青枯雷尔氏菌(李信申等,2016)、小麦蠕孢根腐病菌(张眉等,2016)、杧果炭疽病菌(李江华等,2017)、黄瓜褐斑病菌(吴桥等,2017)和水稻纹枯病菌(张优等,2017)等,而关于淮山炭疽病菌致病力分化研究目前尚无报道。本研究发现来源于我国南方6省(区)的45株淮山炭疽病菌在供试的4个淮山种质材料上的致病力存在明显差异,依据各供试菌株综合致病力表现,分为强、中、弱3种致病类型,以中等致病类型菌株居多,占71.00%,与张眉等(2016)关于山东小麦蠕孢根腐病菌致病力分化、李江华等(2017)关于杧果炭疽病菌致病力分化,以及吴桥等(2017)关于黄瓜褐斑病菌致病力分化的研究结果相似。本研究还发现淮山炭疽病菌致病菌类型的分布比例在不同地理来源问存在明显差异,其中强致病性菌株分布于广西,中致病性菌株分布于贵州、福建和海南,弱致病性菌株分布于四川和云南,表明我国南方淮山炭疽病菌的致病力分化与地理来源有一定的相关性,与李江华等(2017)、吴桥等(2017)的研究结果相似。究其原因,可能与不同省份淮山栽培品种、栽培气候环境、防治用药种类、菌株抗性等因素有一定关联,但具体原因有待进一步探究。
此外,本研究样品的采集地区横跨6省(区),各省份淮山种植区分布广,样品采集的代表性存在一定的局限性,同时各省(区)分离到的菌株数量差异较大,难以准确反映不同来源地菌株的致病力。因此,本研究仅初步探讨了我国南方淮山炭疽病菌的致病力分化情况,要深入了解我国南方淮山炭疽病菌致病力分化的原因,仍需大量采集样品分离菌株,进一步开展相关研究。
4结论
本研究对分离自我国南方6省(区)的45株淮山炭疽病菌进行鉴定,结果表明,45株炭疽病菌菌株均为胶孢炭疽菌(C. gloeosporioide),其在培养特征和系统发育上存在多样性差异,在致病力上存在分化,且受来源地的影响。
(责任编辑 麻小燕)
猜你喜欢
今日农业(2022年12期)2022-07-06 20:15:55
食品科学(2021年17期)2021-09-27 06:47:58
第二课堂(小学版)(2019年7期)2019-07-16 05:26:15
山东化工(2018年23期)2018-12-28 08:01:50
农产品市场周刊(2018年42期)2018-11-08 07:23:46
—— 记福建省安溪县山格淮山专业合作社理事长陈主义
中国农民合作社(2018年8期)2018-09-07 07:32:28
故事会(2018年5期)2018-03-06 22:02:53
农家之友(2018年6期)2018-01-30 21:18:14
农产品市场周刊(2016年4期)2016-05-24 12:57:17
凤凰生活(2015年2期)2015-09-10 07:22:44
