
混种鬼针草属植物对葡萄幼苗生长及镉积累的影响
2018-09-08 03:03:20黄佳璟林立金陈发波廖明安
四川农业大学学报 2018年4期
黄佳璟,林立金,陈发波,王 铤,刘 磊,廖明安*,任 纬
(1.四川农业大学园艺学院,成都 611130;2.四川农业大学果蔬研究所,成都 611130;3.长江师范学院生命科学与技术学院,重庆 408100;4.成都市农林科学院,成都 611130;5.四川省内江市农业科学院玉米研究所,四川内江 641000)
近年来,由于我国工业“三废”的排放以及化肥、农药等化学物质的过量施用,大量重金属进入果园土壤系统,使得果园土壤重金属污染日益严重[1]。其中,镉因其不能在生物体内降解而成为一种极具毒性的重金属元素[2]。葡萄具有丰富的营养物质,外形美观且产品形式多样,是我国重要的果品之一。目前,我国葡萄园已面临着重金属镉污染的危害。研究表明一些地区葡萄园土壤中的镉含量已超过绿色食品产地土壤环境质量标准或镉污染已达到轻度污染等级[3-5]。因此,为了确保葡萄果品的安全、绿色生产,葡萄园镉污染修复显得尤为重要。
植物修复技术普遍被认为具有物理、化学修复方法所无法比拟的成本低廉、安全美观、对环境扰动少等优点,具有广阔的应用前景[6-8]。但目前该方法仍存在如植物生长慢,长势弱,抗性低等缺点,因而限制了植物修复技术优势的充分发挥[9-10]。在农业生产上,混种可提高作物对土壤养分、水、光等资源的有效利用,从而提高其产量与品质[11]。S.N.Whiting等[12] 蓂
将锌超富集植物天蓝遏蓝菜和普通植物菥 混种后发现,超富集植物的锌积累量显著提高,而普通植物的锌积累量却明显降低。有研究发现[13],与大麦混种使得豌豆地上部的Cu、Pb、Zn、Cd和Fe浓度是分别是单种的1.5、1.8、1.4、1.4和1.3倍。研究表明[14],深根的Cd/Zn富集植物柳树(Salix)和矮小的超富集植物拟南芥(Arabidopsis halleri)混种并未增加植物对Cd和Zn的提取效率。由此可见,只有通过合理的混种,才能提高富集植物对重金属的积累,降低果树对重金属的吸收,有效地将植物修复技术应用于果园土壤重金属污染修复。一些研究表明[15-17],鬼针草属(Bidens L.)植物对重金属具有一定的富集特性,且生物量较大、抗性强,可作为先锋植物去除土壤中的重金属污染。本研究通过将4种鬼针草属植物(婆婆针、三叶鬼针草、小花鬼针草、金盏银盘)分别与葡萄幼苗混种,以期筛选出能显著降低葡萄幼苗镉含量的鬼针草属植物,同时为葡萄园镉污染修复提供一定的参考。
1 材料和方法
1.1 供试材料及土壤
供试鬼针草属植物种子采集于四川农业大学成都校区(东经 103°50′,北纬 30°42′)周围农田,鬼针草属植物包括婆婆针(Bidens bipinnata L.)、三叶鬼针草(Bidens pilosa L.)、小花鬼针草(Bidens parviflora Willd.)、金盏银盘(Bidens biternata Merr.et Scherff.)4种;供试葡萄为巨峰的扦插苗,购置于四川省成都市龙泉驿区苗木基地。供试土壤取自四川农业大学成都校区周边农田,其基本理化特性为:pH值7.09,全氮1.50 g/kg,全磷0.76 g/kg,全钾18.02 g/kg,碱解氮 94.82 mg/kg,速效磷 6.30 mg/kg,速效钾149.59 mg/kg,其中供试土壤背景有效镉含量未检出。
1.2 试验设计
试验于2016年4—7月在四川农业大学成都校区进行。2016年4月,将风干后的供试土壤过6.72 mm(3目)筛用21 cm×20 cm(直径×高)塑料盆每盆装入3.0 kg,以CdCl2·2.5H2O分析纯形式加入重金属镉并混匀,使土壤镉浓度为5 mg/kg。保持土壤湿润,自然放置平衡4周,不定期翻土混合,保证土壤充分混合均匀。2016年5月挑选生长基本一致的巨峰葡萄当年扦插幼苗(新梢长约15 cm)及生长一致的鬼针草属植物幼苗(一对真叶展开)栽入塑料盆中,葡萄单种每盆3株,葡萄与超富集植物混种为每盆葡萄2株,鬼针草属植物2株,每个处理重复3次。盆与盆之间的距离为15 cm,完全随机摆放。保证其在自然状况下生长,根据盆中土壤缺水情况,不定期浇水(水中未检出镉),使土壤含水量经常保持在田间持水量的80%左右。为防止污染物淋溶渗漏损失,在盆下放置塑料托盘并将渗漏液倒回盆中。在整个生长过程中不定期的交换盆与盆的位置以减弱边际效应的影响,并及时去除杂草,防治病虫害。
60 d后,选取葡萄植株采集其从上往下的第3或第4片功能叶,测定光合色素(叶绿素a、叶绿素b和类胡萝卜素)含量及抗氧化酶(SOD、CAT)活性,其中光合素色含量采用紫外分光光度法进行测定[18]55-56,SOD活性采用氮蓝四唑法进行测定[18]124-126,CAT活性采用高锰酸钾滴定法进行测定[18]75-76。之后将葡萄和鬼针草属植物整株收获,土壤分开存放。将葡萄植株分成根、茎、叶3部分,将鬼针草属植物植株分为根系和地上部两部分,分别用自来水洗净,再用去离子水冲洗3次后,于110℃杀青15 min,80℃烘干至恒重,称重,然后粉碎,过100目筛,植物样品由硝酸和高氯酸(体积比为4∶1)消化后用iCAP 6300型 ICP光谱仪测定(Thermo Scientific,USA)镉含量[19]。土壤自然风干后过1mm筛,土壤pH值采用土水比1∶2.5进行测定,土壤有效态镉含量由DTPATEA提取并用ICP-MS分析测定[20]。
1.3 数据分析
数据采用SPSS17.0系统进行方差分析(Duncan新复极差法进行多重比较)。根冠比=根系生物量/地上部分生物量,转运系数(TF)=植物地上部分镉含量/根系镉含量[21]。
2 结果与分析
2.1 葡萄幼苗的生物量
从表1可知,与鬼针草属植物混种的葡萄幼苗的根系、茎秆、叶片及地上部分生物量均低于单种。葡萄幼苗的根系及地上部分的生物量均表现为:单种>混种婆婆针>混种三叶鬼针草>混种小花鬼针草>混种金盏银盘,与婆婆针、三叶鬼针草、小花鬼针草和金盏银盘混种的葡萄幼苗的根系生物量分别较单种降低了 3.32%、8.53%、12.80%和 38.39(P<0.05),葡萄幼苗地上部分生物量分别较单种降低了5.01%、9.35%、24.93%和 29.13%(P<0.05)。就根冠比而言,除与金盏银盘混种外,与其他3种鬼针草属植物混种的葡萄幼苗的根冠比均表现为混种高于单种。

表1 混种四种鬼针草属植物对葡萄幼苗生物量的影响Table1 Effects on biomass of grape seedlings by intercropping with four species of Bidens L.
2.2 葡萄幼苗的光合色素含量
由表2可知,与鬼针草属植物混种的葡萄幼苗叶片的叶绿素a、叶绿素b、总叶绿素及类胡萝卜素含量均低于单种。葡萄幼苗叶片的叶绿素a、叶绿素b、总叶绿素总及类胡萝卜素含量均表现为:单种>混种婆婆针>混种三叶鬼针草>混种小花鬼针草>混种金盏银盘。与婆婆针、三叶鬼针草、小花鬼针草和金盏银盘混种的葡萄幼苗叶片的叶绿素a含量较单种分别降低了4.11%、6.93%、9.42%和12.99%(P<0.05),葡萄幼苗叶片的叶绿素b含量较单种分别降低了 4.58%、6.02%、12.61%和 24.36%(P<0.05),葡萄幼苗叶片的类胡萝卜素含量较单种分别降低了5.24%、6.54%、8.64%、和10.21%(P<0.05)。就葡萄幼苗叶片的叶绿素a/b来看,除混种金盏银盘的葡萄幼苗的叶绿素a/b的值达到了3.045之外,混种其他3种鬼针草属植物及单种的葡萄幼苗的叶绿素a/b的值之间差异很小,均在2.700左右。
2.3 葡萄幼苗的抗氧化酶活性
在镉胁迫条件下,混种鬼针草属植物对葡萄幼苗叶片的抗氧化酶(SOD、CAT)活性具有提高或降低作用。从图1可知,混种4种鬼针草属植物均使葡萄幼苗叶片SOD活性显著高于单种(P<0.05),与婆婆针、三叶鬼针草、小花鬼针草和金盏银盘混种的葡萄幼苗叶片SOD活性分别为单种的2.05倍、2.09倍、1.76倍和2.01倍。由图2可知,混种不同的鬼针草属植物对葡萄幼苗叶片的CAT活性影响不同。与婆婆针、三叶鬼针草和小花鬼针草混种使得葡萄幼苗叶片CAT活性降低,分别较单种降低了8.26%、23.91%和 30.43%(P<0.05),与金盏银盘混种使得葡萄幼苗叶片CAT活性提高,较单种提高了18.70%(P<0.05)。
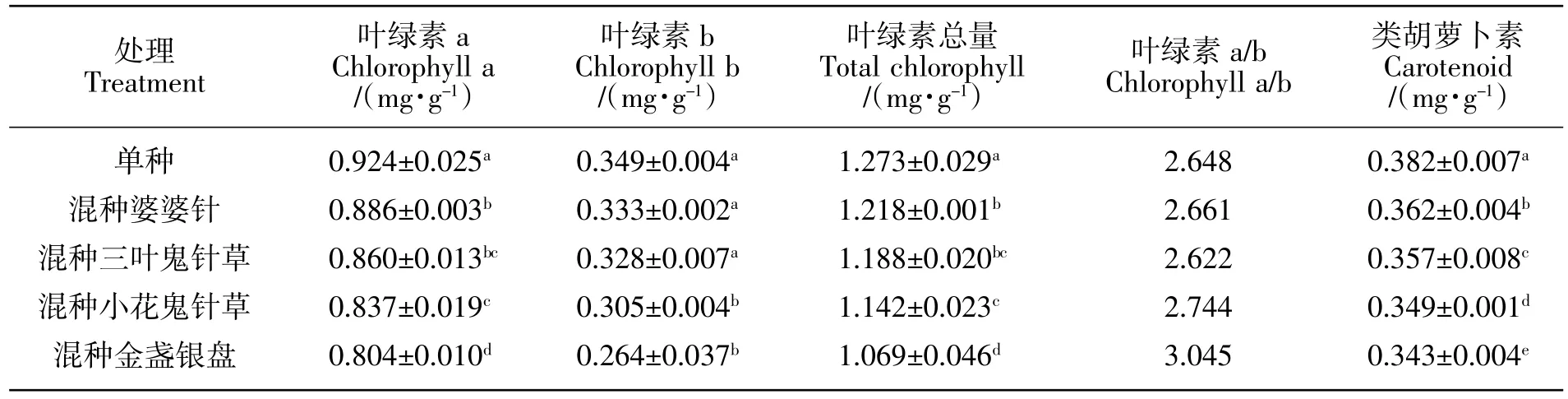
表2 混种四种鬼针草属植物对葡萄幼苗光合色素含量的影响Table2 Effects on photosynthetic pigment content of grape seedlings by intercropping with four species of Bidens L.

图1 葡萄幼苗的SOD活性Figure1 SOD activity of grape seedlings
2.4 葡萄幼苗的镉含量

图2 葡萄幼苗的CAT活性Figure2 CAT activity of grape seedlings
由表3可见,与鬼针草属植物混种的葡萄幼苗的根系、茎秆、叶片及地上部分镉含量均低于单种。葡萄幼苗的根系、茎秆、叶片及地上部分镉含量均表现为:单种>混种婆婆针>混种金盏银盘>混种小花鬼针草>混种三叶鬼针草。按镉含量由高到低,混种的葡萄幼苗的根系镉含量分别较单种降低了5.48%、39.88%、43.46%和 50.72(P<0.05);混种的葡萄幼苗的地上部分镉含量分别较单种降低了13.42%、19.05%、21.86%和 92.21(P<0.05)。就转运系数而言,混种婆婆针和三叶鬼针草降低了葡萄幼苗的转运系数,而混种小花鬼针草和金盏银盘则提高了葡萄幼苗的转运系数。转运系数大小顺序为:混种小花鬼针草>混种金盏银盘>单种>混种婆婆针>混种三叶鬼针草。
2.5 鬼针草属植物的生物量及镉积累
由表4可知,与葡萄幼苗混种的鬼针草属植物的根系及地上部分生物量均表现为:金盏银盘>小花鬼针草>三叶鬼针草>婆婆针,与葡萄幼苗混种的鬼针草属植物的根系镉含量表现为:三叶鬼针草>婆婆针>小花鬼针草>金盏银盘,地上部分镉含量表现为:三叶鬼针草>小花鬼针草>金盏银盘>婆婆针。与葡萄幼苗混种的鬼针草属植物的根系镉积累量表现为:小花鬼针草>三叶鬼针草>婆婆针>金盏银盘,地上部分镉积累量表现为:小花鬼针草>金盏银盘>三叶鬼针草>婆婆针。
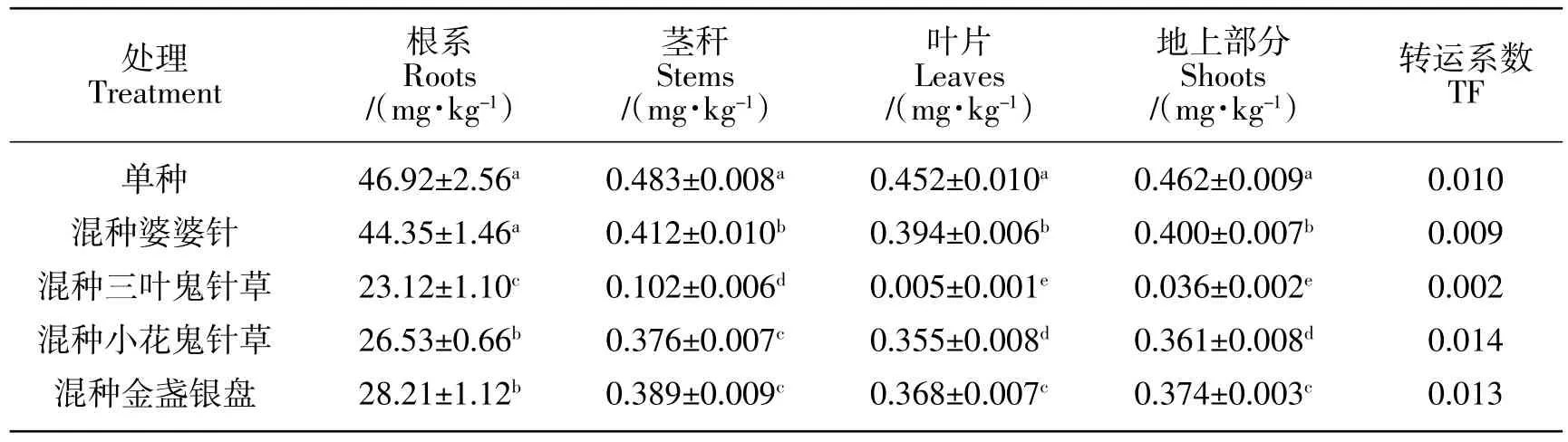
表3 混种4种鬼针草属植物对葡萄幼苗镉含量的影响Table3 Effects on cadmium content in grape seedlings by intercropping with four species of Bidens L.

表4 与葡萄混种的四种鬼针草属植物生物量及镉积累Table4 The biomass and cadmium accumulation of four species of Bidens L.by intercropping with grape seedlings
2.6 混种后土壤pH值及有效态镉含量
由图3可知,各混种处理后土壤的pH值均显著低于单种(P<0.05),具体表现为:单种>婆婆针混种葡萄>三叶鬼针草混种葡萄>小花鬼针草混种葡萄>金盏银盘混种葡萄。就土壤有效态镉含量而言(见图4),除了金盏银盘与葡萄混种后的土壤有效态镉含量高于单种外,其余3种混种处理后土壤的有效态镉含量均显著低于单种(P<0.05),具体表现为:单种>婆婆针混种葡萄>小花鬼针草混种葡萄>三叶鬼针草混种葡萄,与婆婆针、三叶鬼针草和小花鬼针草混种后土壤的有效态镉含量分别较单种降低了1.38%、4.84%和4.61%。

图3 土壤pH值Figure3 Soil pH value
3 讨论

图4 土壤有效态镉含量Figure4 Soil available cadmium concentration
在重金属污染条件下,混种的植物通过根系分泌不同的有机物改变土壤根际环境状况,从而可以降低或升高土壤中重金属的生物有效性,促进或抑制植物的生长及生理生化过程[22-24]。所以,合理的搭配混种植物,才能得到促进植物生长及减少重金属吸收的最佳方式。混种对于重金属胁迫条件下不同种类植物的生物量的影响存在3种情况:一是使一方生物量增加,另一方生物量减少;二是使双方的生物量均增加;三是对双方生物量均无明显影响[25]。本研究中,由于混种的4种鬼针草属植物均为农田杂草,生长速度及生存竞争力均高于葡萄幼苗,故本研究结果表现为混种的葡萄幼苗生物量均低于单种,即混种使得葡萄一方的生物量降低,这与前人对樱桃混种富集植物的研究结果相似[26]。光合作用是植物最重要的生理反应之一,其光合色素含量水平高低直接反映了光合能力的强弱。本研究中,与鬼针草属植物混种的葡萄幼苗叶片的叶绿素a含量、叶绿素b含量、叶绿素总含量及类胡萝卜素含量均低于单种,与葡萄幼苗生物量的表现一致,表明光合素色含量同植物的生长表现基本一致,促进植物生长的同时一般会伴随光合色素含量水平的提高,而生物量的减少一般也会伴随着植物光合色素含量水平的降低。重金属胁迫可导致植物体改变自身防御机制从而免受活性氧自由基的损伤,其中活性氧自由基的清除是一种普遍的防御机制,它通过抗氧化酶系统中的重要酶类(如SOD、CAT)进行保护防御[27]。本研究中,在重金属镉胁迫下,与4种鬼针草属植物混种提高了葡萄幼苗叶片的SOD活性,这与对铅锌胁迫下混种三叶草对假繁缕的研究结果一致[28]。另外,本研究除与金盏银盘混种提高了葡萄幼苗叶片的CAT活性,与其他3种鬼针草属植物混种则降低了葡萄幼苗叶片的CAT活性,这与对铅锌胁迫下混种黑麦草对假繁缕的研究结果基本一致[28]。表明混种鬼针草属植物对葡萄幼苗的各抗氧化酶影响不同,可能是由于不同抗氧化酶对重金属的敏感度不同[29]。
不同植物混种后,由于各自对养分和重金属的吸收能力与抗性的不同及根系分泌物的差异,会导致根际土壤环境的改变,进而表现为促进或抑制植株生长和重金属吸收[24]。本研究中,混种4种鬼针草属植物使得葡萄幼苗的根系及地上部分的镉含量均低于单种,表明混种富集植物可以降低普通植物的镉含量,这与前人的一些研究结果一致[26,30]。本研究的葡萄幼苗根系及地上部分镉含量表现为:单种>混种婆婆针>混种金盏银盘>混种小花鬼针草>混种三叶鬼针草,结合生物量来看,葡萄根系及地上部分生物量表现为:单种>混种婆婆针>混种三叶鬼针草>混种小花鬼针草>混种金盏银盘,生物量是混种均低于单种不利于植物的生长,但混种后葡萄幼苗特别是地上部分的镉含量显著降低有利于降低植物的镉积累从而用于植物的镉修复的生产应用,特别是有利于果品的安全绿色生产。其中与三叶鬼针草混种的葡萄幼苗镉含量显著低于其他处理及单种,且生物量下降幅度相对较小,故综合比较,三叶鬼针草混种葡萄最有利于葡萄的生长及镉修复的生产应用。另外,与葡萄混种后,4种鬼针草属植物地上部分镉含量表现为:三叶鬼针草>小花鬼针草>金盏银盘>婆婆针。表明2种植物混种后,可能会增加其中一种植物的重金属积累量,而减少另一种植物对重金属的吸收[12,31],本研究即表现为葡萄镉吸收减少,鬼针草属植物镉吸收增加。重金属污染下,根际环境是影响植物提取土壤重金属的主要因素,而混种产生的“根际对话”则可影响植物及土壤环境的重金属含量水平[32-33]。本研究中,混种后土壤的pH值显著低于单种,表明混种鬼针草属植物会使得葡萄园土壤酸性增强,此改变趋势与前人关于混种小飞蓬对酸樱桃的土壤pH值的改变趋势相同[34]。对于土壤的有效态镉含量,除金盏银盘与葡萄混种外,其余3种混种处理后土壤的有效态镉含量均低于单种,这与前人在混种富集植物对樱桃及酸樱桃的土壤有效态镉含量的结果不一致,可能是由于除土壤pH值会影响土壤的镉的有效性及固化外,还存在其他影响因素[26,34]。其中三叶鬼针草混种葡萄后土壤有效态镉含量最低,表明混种某些鬼针草属植物可降低土壤的镉含量,减少环境中镉的影响,从而有助于改善土壤环境状况。由此可见,本研究与葡萄混种的4种鬼针草属植物中,三叶鬼针草使葡萄幼苗镉含量降低最多且生物量不大幅下降,故三叶鬼针草是能显著降低葡萄幼苗镉含量,用于葡萄园土壤镉修复的最佳材料。
4 结论
混种4种鬼针草属植物(婆婆针、三叶鬼针草、小花鬼针草、金盏银盘)降低了葡萄幼苗生物量、光合色素含量及除与金盏银盘混种的葡萄幼苗的CAT活性,但提高了混种的葡萄幼苗的SOD活性。混种4种鬼针草属植物使得葡萄幼苗的镉含量均低于单种,其中与三叶鬼针草混种的葡萄幼苗镉含量最低,显著低于与其他3种鬼针草属植物混种的及单种的葡萄幼苗镉含量。对于土壤有效态镉含量,除了金盏银盘与葡萄混种后的土壤有效态镉含量高于单种外,其余3种混种处理后土壤的有效态镉含量均低于单种,其中三叶鬼针草混种葡萄后土壤有效态镉含量最低。因此,本研究的4种鬼针草属植物均可降低葡萄幼苗的镉含量,可应用于葡萄园镉污染土壤的修复,且最佳材料是三叶鬼针草。
猜你喜欢
食品研究与开发(2021年19期)2021-11-17 06:59:10
雪豆月读·低年级(2021年1期)2021-09-10 07:22:44
河北画报(2020年19期)2020-06-16 13:36:22
大理文化(2020年3期)2020-06-11 00:41:51
读写月报(初中版)(2019年9期)2019-09-10 07:22:44
快乐语文(2018年34期)2018-03-06 09:18:22
江苏农业科学(2017年24期)2018-01-12 11:39:25
环境科技(2017年1期)2017-09-05 03:08:02
农业与技术(2017年6期)2017-04-21 00:01:27
大自然探索(2015年5期)2015-09-10 07:22:44
