
基于SSR标记的杂交大豆主要亲本遗传多样性分析
2018-09-07 11:09白志元张瑞军武国平连世超杨玉花卫保国
华北农学报 2018年4期
白志元,张瑞军,武国平,连世超,杨玉花,卫保国
(1.山西省农业科学院 农作物品种资源研究所,农业部黄土高原作物基因资源与种质创制重点实验室,杂粮种质资源发掘与遗传改良山西省重点实验室,山西 太原 030031;2.山西省农业科学院 园艺研究所,山西 太原 030031)
大豆高产育种一直以来都是育种工作者的重要目标之一。杂种优势现象在生物界普遍存在,利用杂种优势成为提高社会经济效益必不可少的工具[1]。水稻、玉米、谷子、高粱等作物已利用杂种优势大幅度提高了产量,在主要作物中,大豆杂种优势利用研究相对滞后[2]。大豆是严格的自花授粉作物,天然异交率不到1%。随着大豆细胞质雄性不育系的发现,大豆杂种优势利用成为可能[3-4],该项研究对提升我国大豆产业具有战略性意义。目前,我国在该领域处于世界领先水平,国内多家单位已实现了杂交大豆生产的三系配套体系,其中,吉林省农业科学院、安徽省农业科学院、安徽省阜阳市农业科学院和山西省农业科学院已审定了杂交大豆品种[5-8]。
杂交大豆产业化有2项瓶颈技术:高效繁种制种技术和强优势组合的筛选。山西省农业科学院农作物品种资源研究所杂交大豆课题组对这2项技术进行了有效的攻关,形成了杂交大豆生产所需的规范繁种制种技术[9],制定了《大豆杂交种制种技术规程》、《大豆杂交种种子》山西省地方标准;审定了山西省首个杂交大豆品种-晋豆48号[8]。然而,本课题组在高产组合的筛选上仍不理想,没有明显突破。大豆杂交育种研究表明,亲本的亲缘关系对大豆杂种优势有着重要的影响[10-11]。遗传多样性能反映品种之间亲缘关系的远近,能对作物的育种应用提供重要的参考依据。许多研究表明,SSR标记具有简便快速、重复性高和多态性丰富等优点,并具有很好的稳定性,在大豆遗传多样性研究中已得到广泛应用[12-16]。
本研究选用64对覆盖大豆20条染色体的SSR标记,对本课题组现有主要的32份杂交大豆亲本(包括15份保持系与17份恢复系)进行遗传多样性和亲缘关系的分析,以期为山西三系杂交大豆育种中保持系与恢复系的筛选、遗传改良以及强优势组合的选育等方面提供参考。
1 材料和方法
1.1 试验材料
选用本课题组自育的14个不育系对应的保持系和南京农业大学选育的NJCMS3A对应的保持系NJCMS3B,共15份材料;恢复系为17份材料,具体名称如表1所示。
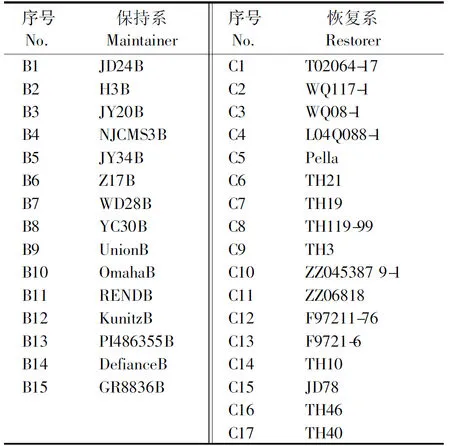
表1 供试保持系与恢复系名称Tab.1 The name of the maintainer and restorer of the test
1.2 基因组DNA的提取
取大豆幼嫩叶片,在液氮中研磨后,采用CTAB[17]法提取叶片DNA,并检测其浓度和质量。
1.3 PCR扩增与产物检测
选用分布在大豆20条染色体上的64对SSR引物进行分析,该引物由生工生物工程(上海)股份有限公司合成(表2)。PCR反应采用10 μL体系:2.5 mmol/L dNTP 0.5 μL,10 μmol/L Primer 2 μL,10×PCR Buffer 1 μL,5 U/μLTaqPolymerase 0.1 μL,10 ng/μL模板 DNA 5 μL,ddH2O 1.4 μL。PCR反应在美国Bio-Bad 1000-系列PCR仪上进行,反应条件为:94 ℃预变性5 min;94 ℃变性1 min,优化的退火温度(依引物退火温度而定)退火30 s,72 ℃延伸30 s,34 个循环;72 ℃温育10 min;4 ℃保存。扩增产物经8%非变性聚丙烯酰胺凝胶电泳、银染、拍照记录试验结果。
1.4 数据的统计分析
根据每对SSR产物的银染结果统计数据,有扩增条带表示为“1”,无条带者表示为“0”。材料间的遗传相似系数(Genetic similarity,GS)按 Nei等[18]的方法计算,遗传相似性聚类按遗传相似系数的值以不加权成对算术平均法(UPGMA)进行聚类,利用NTSYS-PC2.1软件绘制聚类图。
多态性信息量(Polymorphism information content)PIC计算公式如下。
式中,Pij表示位点i的第j个等位变异出现的频率[19],PIC值位于0~1,其中,0代表该位点无多态性,1代表有较多的等位位点。
2 结果与分析
2.1 SSR标记的多态性分析
本研究用分布于大豆20条染色体的64对SSR标记对32份杂交大豆亲本(包括15份保持系与17份恢复系)进行多态性分析。从表2可以看出,试验共检测出357个等位基因变异,单个位点检测到的等位基因数变化为2~12个,平均为5.578个。其中,Satt358、Satt380这2对引物等位位点最多,均为12个,引物Satt211等位位点最少,为2个。SSR位点的多态性信息指数PIC变幅为0.368~0.900,平均为0.666。
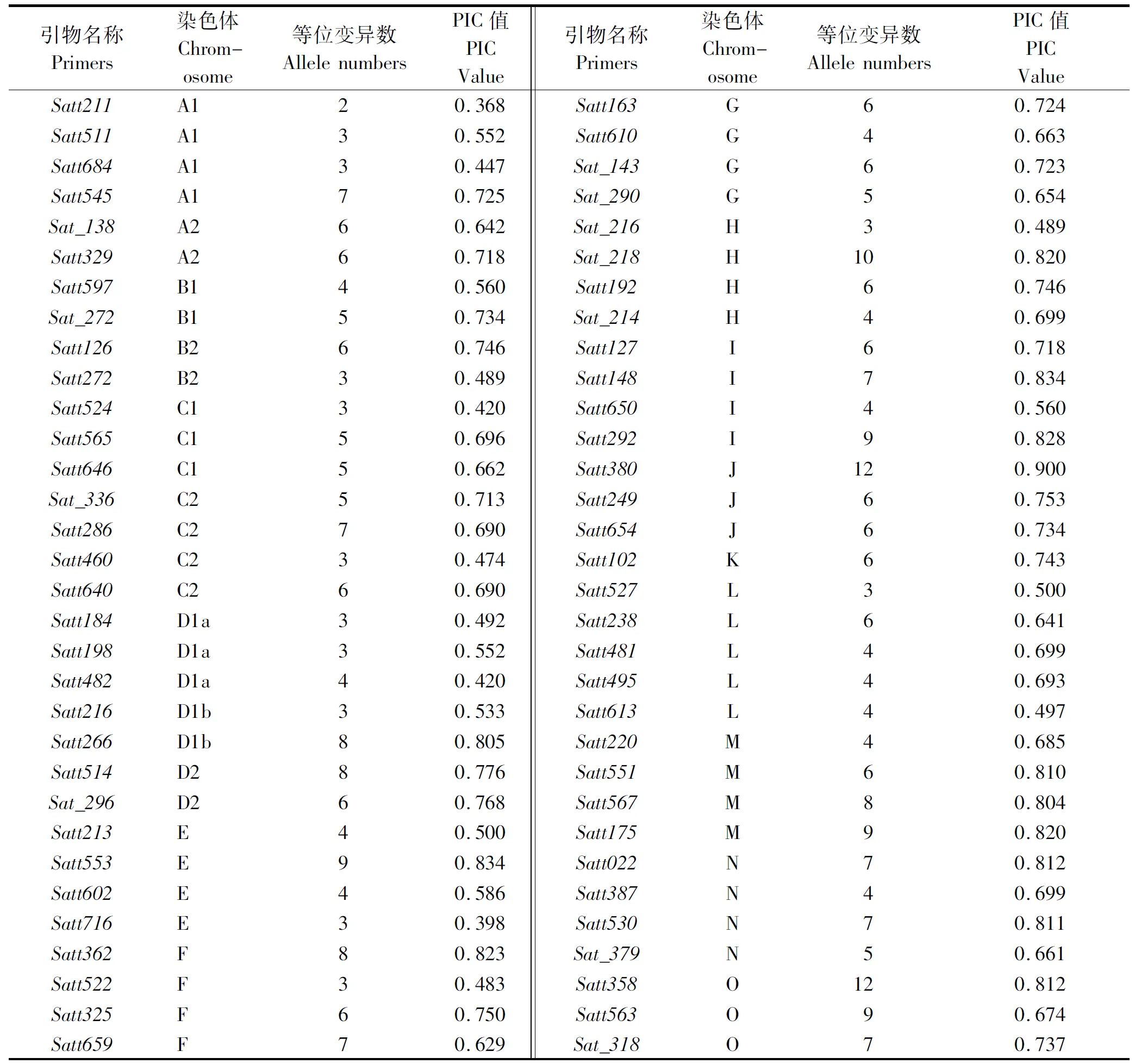
表2 64对SSR引物及所在染色体、等位变异数和PIC值Tab.2 The chromosome loci amplified allele number and PIC value of 64 SSR primers
2.2 保持系与恢复系SSR标记的多态性分析
从表3可以看出,用分布于大豆20条染色体的64对SSR标记进行扩增,15份保持系共检测出322个等位变异,单个位点检测到的等位基因数变化为2~9个,平均为5.031个,其中,Satt358、Satt380、Satt292这3对引物等位位点最多,均为9个,Satt211、Satt213、Satt527这3对引物等位位点最少,均为2个;SSR位点的多态性信息含量(PIC)变幅为0.370~0.868,平均为0.661,以引物Satt380最高,引物Satt211最低。17份恢复系共检测出320个等位变异,单个位点检测到的等位基因数变化为2~12个,平均为5个,其中,引物Satt380等位位点最多,为12个;SSR位点的多态性信息含量(PIC)变幅为0.329~0.904,平均为0.630,以引物Satt380最高,引物Satt460最低。

表3 64对SSR引物分别在15份保持系、17份恢复系中等位变异数和PIC值Tab.3The amplified allele number and PIC value of 64 SSR primers in 15 maintainers and 17 restorers
2.3 保持系间的遗传差异与遗传关系
根据SSR 统计数据计算保持系间的遗传相似系数(GS),GS值变幅为0.491~0.857,平均为0.637。其中,B4(NJCMS3B)和B13(PI486355B)的遗传相似系数最小,为0.491;B12(KunitzB)和B15(GR8836B)的遗传相似系数最大,为0.857。
利用NTSYS-PC2.1软件通过UPGMA法进行聚类分析,以GS值0.61为标准,15份保持系共聚成两类(图1)。其中,第Ⅰ类包括B1(JD24B)、B2(H3B)、B3(JY20B)、B5(JY34B)和B13(PI486355B)共5个保持系,其余10个保持系被聚在第Ⅱ类。在相似系数0.65处,第Ⅱ类又可分为3个亚类,其中,第Ⅰ亚类含有B4(NJCMS3B)和B8(YC30B)共2个保持系,第Ⅱ亚类包括B9(UnionB)、B10(OmahaB)、B12(KunitzB)、B15(GR8836B)、B14(DefianceB)和B11(RENDB)共6个保持系,第Ⅲ亚类含有B6(Z17B)和B7(WD28B)共2个保持系。
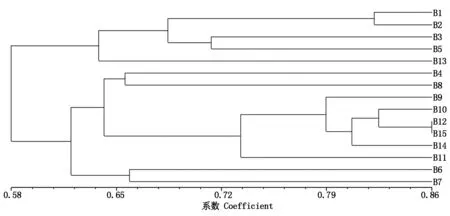
图中序号与表1中名称对应。图2同。The name of the sequence number in the graph is shown in Tab.1. The same as Fig.2.
2.4 恢复系间的遗传差异与遗传关系
根据SSR统计数据计算恢复系间的遗传相似系数(GS),GS值变幅为0.494~0.969,平均为0.669。其中,C15(JD78)和C17(TH40)的遗传相似系数最小,为0.494;C6(TH21)和C7(TH19)的遗传相似系数最大,为0.969。
利用NTSYS-PC2.1软件通过UPGMA法进行聚类分析,以GS值0.65为标准,17份恢复系共聚成两类(图2)。其中,第Ⅰ类包括C1(T02064-17)、C3(WQ08-1)、C2(WQ117-1)、C4(L04Q088-1)、C8(TH119-99)、C9(TH3)、C10(ZZ045387 9-1)、C5(Pella)、C15(JD78)、C6(TH21)、C7(TH19)和C11(ZZ06818)等12个恢复系,其余5个恢复系被聚在第Ⅱ类,分别为C16(TH46)、C17(TH40)、C12(F97211-76)、C13(F9721-6)和C14(TH10)。

图2 17份恢复系的遗传多样性聚类结果Fig.2 Dendrogram of cluster analysis based on genetic similarity coefficient between 17 restorers
3 讨论
SSR分子标记是分子辅助育种、材料基因型鉴定、种属间亲缘关系和遗传多样性等研究的有力工具。王彩洁等[13]对中国自20世纪40年代以来大豆主产区东北和黄淮海地区大面积种植的89个大豆品种进行遗传多样性分析,结果显示,平均等位变异数为5.48个,等位变异的变化为1~14个,黑龙江北部、黑龙江中南部、吉林辽宁地区和黄淮海地区大面积种植品种标记的多态性信息含量(PIC)依次为0.414,0.469,0.522和0.562。Wang等[20]研究结果显示,40份山西大豆育成品种、地方品种和野生资源平均位点的等位变异数为6.55,PIC为0.585~0.850,平均为0.78。Diwan等[21]对北美大豆品种的35个祖先亲本多样性进行分析,结果显示,平均每个SSR引物的等位变异数为10.1,PIC为0.64~0.91,平均为0.8。本研究64对引物对32份杂交大豆亲本材料共检测出357个等位基因变异,单个位点检测到的等位基因数变化为2~12个,平均为5.578个,位点多态性信息含量(PIC)变幅为0.368~0.900,平均为0.666。本研究多样性信息含量结果介于中外学者之间,主要是由于本试验材料除黄淮与东北材料外,还有一些美国材料。上述结果也进一步证明,SSR标记检测是研究大豆品种资源遗传差异的有效手段之一。
崔艳华等[14]对96份黄淮夏大豆进行遗传多样性分析,聚类结果表明,96份材料呈现一定的地理分布规律。王彩洁等[13]研究表明,同一区域内大面积种植品种的同质化现象相当明显。本研究在聚类分析中也发现,聚类结果与地理来源具有一定相关性,地理来源相近的材料聚为一类,如保持系聚类中第Ⅱ类中第Ⅱ亚类的6个保持系皆来自于美国。此外,对系谱来源不清或者血缘不直接相关的种质划分到相应的类群,如保持系第Ⅰ类中B2(H3B)和B13(PI486355B);恢复系第Ⅰ类中C9(TH3)、C6(TH21),第Ⅱ类中C16(TH46)等材料,这将对保持系、恢复系材料的利用、种质创制及从地理来源有目的地筛选保持系与恢复系具有很现实的指导意义。此外,本研究保持系的多态性信息含量(PIC)0.661高于恢复系的0.630,保持系间的平均遗传相似系数(GS)0.637低于恢复系的0.669,这也与笔者在筛选保持系和恢复系的结果一致,保持材料比恢复材料容易找到。
多年来,山西杂交大豆育成的组合及品种在单产、抗性等方面未取得较明显突破,其重要原因之一在于不育系、保持系及恢复系遗传基础狭窄。近年来,山西省农业科学院农作物品种资源研究所杂交大豆课题组在大豆种质资源的扩增改良方面取得了一定的成效,拓宽了杂交大豆的亲本材料,但种质基础仍相对狭窄。因此,在今后的育种工作中,应继续从全世界种质资源中筛选不育系、保持系及恢复系种质作为研究的基础材料;同时,还须加大利用资源,拓宽杂交大豆不育系、保持系及恢复系的遗传基础,从而实现山西杂交大豆育种的新突破。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
作物学报(2022年6期)2022-04-08
昆明医科大学学报(2022年2期)2022-03-29
园艺与种苗(2021年10期)2021-11-28
国际医学放射学杂志(2021年5期)2021-10-22
昆明医科大学学报(2021年6期)2021-07-31
昆明医科大学学报(2021年3期)2021-07-22
粮食科技与经济(2020年7期)2020-10-20
中国种业(2018年8期)2018-08-28
麦类作物学报(2018年4期)2018-05-11
